101. The Significance of the Skeleton.-The skeleton of the Radiolaria is developed in such exceedingly manifold and various shapes, and exhibits at the same time such wonderful regularity and delicacy in its adjustments, that in both these respects the present group of Protista excels all other classes of the organic world. For, in spite of the fact that the Radiolarian organism always remains merely a single cell, it shows the potentiality of the highest complexity to which the process of skeleton formation can be brought by a single cell. All that has been brought to pass in this direction by single tissue-cells of animals and plants does not attain the extremely high stage of development of the Radiolaria. Only very few Rhizopoda of this very rich and varied class fail to exhibit the power of forming this firm supporting and protecting organ-indeed, only ten of the seven hundred and thirty-nine genera which are enrolled in the list of the Challenger collection, namely, six genera of SPUMELLARIA (five Thalassicollida, Actissa, Thalassolampe, Thalassopila, Thalassicolla, Thalassophysa, Pl. 1, and one genus of Collozoida, Collozoum, Pl. 3), and in addition two genera of NASSELLARIA (the Nassellida, Cystidium and Nassella, Pl. 91, fig. 1), and two genera of PHAEODARIA (the Phaeodinida, Phaeocolla and Phaeodina, Pl. 101, figs. 1. 2). These skeletonless forms of Radiolaria are, however, of extreme interest, since they include the original stem-forms of the whole class as well as of its four legions. All Radiolaria which form skeletons have originated from soft and skeletonless stem-forms by adaptation, and that polyphyletically, for the skeletal types of the four legions have been developed independently of each other (§ 108).
102. The Chemical Peculiarities of theSkeleton.-The chemical composition of the skeleton shows very marked variations in the different legions of the Radiolaria. The two legions SPUMELLARIA and NASSELLARIA (united formerly as "Polycystina") form their skeleton of pure silica (see note A, below) ; the legion PHAEODARIA of a silicate of carbon (see note B), and the ACANTHARIA of a peculiar organic substance-acanthin (see note C). This explains the well-known fact that the deposits of fossil Radiolaria (or Polycystine marls) are composed exclusively of the skeletons of SPUMELLARIA and NASSELLARIA, those of the ACANTHARIA and PHAEODARA being entirely absent (in the case of the last group, however, exception must be made in favour of the Dictyochida, or those PHAEODARIA whose skeleton is made up of isolated scattered tangential siliceous fragments). The enormous deposits of Radiolarian skeletons in the deep sea of today, which constitute the Radiolarian ooze, consist, like the fossil Polycystine marls, almost exclusively of the shells of SPUMELLARIA and NASSELLARIA, though here the acanthin skeletons of the ACANTHARIA may be present in very small numbers, and the silicate skeletons of the PHAEODARIA, which offer more resistance to the solvent action of sea-water, somewhat more abundantly. Calcareous skeletons do not occur in the Radiolaria (see note D).
A. The pure siliceous skeletons of the Polycystina were first recognised in 1838 by Ehrenberg in chalky marls (L. N. 2, p. 117). Since the two legions ACANTHARIA and PHAEODARIA were entirely unknown to Ehrenberg, his name Polycystina has reference only to the SPUMELLARIA and NASSELLARIA.
B. The silicate skeleton of the PHAEODARIA was formerly taken by me for a purely siliceous one. When I described the first PHAEODARIA in my Monograph in 1862, I was only acquainted with five genera and seven species, whilst the number of PHAEODARIA here described from the Challenger amounts to eighty-four genera and four hundred and sixty-five species. In the great majority of these (though not in all) the skeleton becomes more or less intensely stained by carmine, and is also more or less charred at a red heat, in some even becoming of a blackish-brown. In many PHAEODARIA, furthermore, the hollow skeletal tubes are destroyed by the continued action of heat. They are also, for the most part, strongly acted upon, or even destroyed by boiling caustic alkalis, whilst boiling mineral acids have no effect upon them. The best method of cleaning the skeletons of PHAEODARIA from their soft parts is to heat them in concentrated sulphuric acid, and then add a drop of fuming nitric acid ; in this they are not dissolved even on prolonged heating. From these facts it would appear that the skeletons of the PHAEODARIA consist of a compound of organic substance and silica, or a "carbonic silicate." The more intimate composition yet remains to be discovered, as also the manifold differences which the various families of PHAEODARIA seem to show in respect of its composition. The small skeletal fragments of the Dictyochida (the only remains of PHAEODARIA which occur as fossils) appear to consist of pure silica.
C. The acanthin skeleton of the ACANTHARIA was first described as such in my Monograph (1862, pp. 30-32). Johannes Müller, the discoverer of this legion, took them for siliceous skeletons and defined the Acanthometra as "Radiolaria without lattice-shell, but with siliceous radial spines" (L. N. 12, p. 46). I formerly supposed that the acanthin skeletons in some of the ACANTHARIA were partially or wholly metamorphosed into siliceous skeletons, but, according to the investigations of R. Hertwig, this does not appear to be the case ; he showed that the skeletons of the most varied Acanthometra and Acanthophracta are completely dissolved under the longer or shorter action of acids, and supposes that in all ACANTHARIA, without exception, the skeleton is composed of acanthin (1879, L. N. 33, p. 120). Quite recently Brandt has found that the acanthin spines dissolve not only in acids, alkalis, and "liquor conservativus" (as I had shown), but also in solutions of carbonate of soda (1 per cent.), and even of common salt (10 to 20 per cent.) ; he concludes from this that they consist of an albuminoid substance (vitellin) (L. N. 38, p. 400). I am unable to share this view, for I have never been able to see some of the most important reactions of albumen in any of the skeletons which I have examined, such for example as the xanthoproteic reaction, the red coloration with Millon's test, &c. They do not become yellow either with nitric acid or with iodine. In dilute mineral acids they dissolve more rapidly than in concentrated. My usual method of cleansing the skeleton of ACANTHARIA (which has been practised with the same result on thousands of specimens) consists in heating the preparation in a small volume of concentrated sulphuric acid and then adding a drop of fuming nitric acid ; all other constituents (the whole central capsule and the calymma) are thus very rapidly destroyed ; the skeleton remains quite uninjured and withstands the combined action of the mineral acids for a longer or shorter time, though on prolonged heating it also is dissolved. I do not therefore regard acanthin as an albuminous substance, but as one related to chitin.
D. Calcareous skeletons have not been certainly demonstrated in the Radiolaria, and probably do not occur. Sir Wyville Thomson in his Atlantic (1877, L. N. 31, vol i. p. 233, fig. 51) described under the name Calcaromma calcarea, a Radiolarian which contained scattered in its calymma numerous calcareous corpuscles "resembling the rowels of spurs." These are identical with the "toothed bodies, recalling crystal balls," which Johannes Müller figured in the Mediterranean Thalassicolla morum so early as 1858, and compared with the "siliceous asterisks of Tethya" (L. N. 12, p. 28, Taf. vii. figs. 1, 2). I formerly regarded these peculiar calcareous corpuscles, whose solubility in mineral acids I had observed, as spicules of a Thalassicollid, and hence described the species in my Monograph as Thalassosphaera morum (L. N. 16, p. 260). I have, however, seen reason to change my view, and am now led to suppose that those peculiar calcareous corpuscles, which may be named " Calcastrella," are not formed by the Radiolarian itself, but are foreign bodies which have been accidentally incorporated into the calymma of a Thalassicollid (Actissa). These corpuscles occur, often in large numbers, in many preparations in the Challenger collection, and in the calymma of other Radiolaria, chiefly Discoidea, hence it would appear that they are foreign bodies taken up by the pseudopodia and carried into the calymma by the circulation of the sarcode. The Radiolaria which Sir Wyville Thomson figured as Calcaroromma calcarea, and Müller as Thalassicolla morum, I regard as species of Actissa (see p. 13), perhaps Actissa radiata of the Pacific, and Actissa primordialis of the Mediterranean (compare the description of the Thalassosphaerida of the Challenger collection, pp. 30, 31).
103. The Physical Properties of the Skeleton.-The skeletons of all Radiolaria are characterised pre-eminently by a high degree of firmness, which fits them to serve as protective and supporting apparatus. This is obvious in the case of the pure siliceous shells of the Polycystina ; but the acanthin framework of the ACANTHARIA also possesses a degree of stiffness but little inferior, whilst the silicate skeletons of the PHAEODARIA seem on the whole to be not so firm. The hollow skeletal tubes of the last-named, which are filled with gelatinous material, are very brittle on account of the delicacy of their walls. Their elasticity also is very small, whilst that of the acanthin spines is considerable. The thin long needles of many ACANTHARIA are very elastic, as are also the bristle-like siliceous spicules of many SPUMELLARIA. The refractive power of the skeleton in the various legions is very different, depending upon the chemical constitution. The siliceous skeleton of the Polycystina (SPUMELLARIA and NASSELLARIA) and the silicate skeleton of the PHAEODARIA have the same refractive index as glycerine, and hence become invisible when mounted in that fluid ; they then become visible only on addition of water, and are clearer in proportion to the quantity of water which is added. The refractive index of acanthin is, however, very different from that of glycerine, so that the skeletons of ACANTHARIA are readily visible when mounted in this fluid. In water, the skeletons of all Radiolaria appear about equally refractive, as also in Canada balsam. The substance of the skeleton appears almost entirely hyaline, colourless, and transparent. Very rarely it is faintly coloured (in some ACANTHARIA). A cloudy opaque constitution is seen in some PHAEODARIA (especially in the "porcellanous shells" of Tuscarorida and Circoporida, Pls. 100, 114, 115, 116, 117); when dried, these appear by reflected light milky-white or yellowish-white ; the cause of this opacity lies partly in the peculiar "cement-like structure" of these porcellanous shells, partly in their fine porosity, and the minute air-bubbles contained in their thick walls.
104. The Elementary Structure of the Skeleton.-The general constitution of the skeletonor more accurately expressed, of the morphological elements of which the skeleton consistsis of such a nature that it may be termed structureless. Both the organic acanthin skeletons of the ACANTHARIA and the silicate skeletons of the PHAEODARIA, as well as the inorganic siliceous skeletons of the SPUMELLARIA and NASSELLARIA, appear under the microscope perfectly homogeneous, transparent, colourless, and crystalline. Only very rarely do they show traces of a concentric striation, which arises from the deposition of the skeletal substance in layers ; as, for example, the thick spines of some PHAEODARIA (Pls. 105, 106, 107, &c.). Some of the PHAEODARIA, however, form an exception to this rule, inasmuch as their partially tubular skeletal elements possess a remarkable porcellanous structure. In the tubular or Cannoid skeleton, which occurs in most CANNOPYLEA, the lumen of the thin-walled flinty tube is filled with jelly, and frequently a thin siliceous thread runs in its axis, and is connected with the wall by transverse threads (§§ 127, 139). The elementary structure of the opaque porcellanous shells, which distinguish the two families Circoporida (Pls. 114, 115, 116, 117) and Tuscarorida (Pl. 100), is quite peculiar. Numerous fine siliceous spicules lie scattered irregularly in a finely granular or porous matrix.
105. Complete and Incomplete Lattice-Shells.-In the great majority of Radiolaria (in all four legions) the skeleton has the form of a delicate lattice-shell or a receptacle in which the central capsule is enclosed. In a small minority, however, this is not the case. The skeleton then consists ouly of isolated rigid pieces (radial or tangential spicules), or of a simple ring (sagittal ring of the Stephoidea), or of a basal tripod with or without a loose tissue of trabeculae, &c. (Plectoidea) ; the central capsule is then not surrounded by a special latticed receptacle, but only rests upon the skeletal trabeculae. According to these different arrangements, two principal groups or sublegions may be distinguished in each legion, of which one set (Cataphracta) are characterised by a complete lattice-shell, whilst the others (Aphracta) are without it. The RADIOLARIA APHRACTA, then, or Radiolaria without a complete skeleton, are the Collodaria (p. 9), the Acanthometra (p. 725), the Plectellaria (p. 895), and the Phaeocystina (p. 1543). On the other hand, the RADIOLARIA CATAPHRACTA, or Radiolaria with a complete skeleton, are the Sphaerellaria (p. 49), the Acanthophracta (p. 791), the Cyrtellaria (p. 1015), and the Phaeocoscina (p. 1590).
Upon this basis the first subdivision of the Radiolaria was made by Johannes Müller, who recognised three groups :-" I. Thalassicolla, without receptacle, naked or with spicules ; II. Polycystina, with a siliceous receptacle ; III. Acanthometra, without receptacle, but with siliceous radial spines" (L. N. 12, p. 16).
106. The Ectolithia and Entolithia (Extracapsular and Intracapsular Skeletons).- The relation of the skeleton to the central capsule in the Radiolaria is very various in many respects ; in the first instance two great groups, Ectolithia and Entolithia(see note A), may be distinguished topographically by mere external observation ; in the former the skeleton lies entirely outside the central capsule ; in the latter, partially at all events, within it. The Ectolithia, with a completely extracapsular skeleton, include all NASSELLARIA and PHAEODARIA, as well as a great part of the SPUMELLARIA (all Collodaria and the most archaic forms of Sphaerellaria) ; the Entolithia, on the other hand, in which the skeleton lies partly within, partly without the central capsule, include all ACANTHARIA and the majority of the SPUMELLARIA (most Sphaerellaria, see note B).
A. The difference between Ectolithia and Entolithia was applied in my Monograph in 1862 (p. 222) to separate the Monocyttaria into two main groups. The arrangement was, however, quite artificial, being contrary to the natural relations of the larger groups, as was shown seventeen years later by the discovery of the different structural relations of the central capsule.
B. Among the ACANTHARIA, which all possess primitively an intracapsular and centrogenous skeleton, the remarkable Cenocapsa (Pl 133, fig. 11), seems to furnish the single exception ; in it the skeleton consists of a simple spherical shell which encloses the concentric central capsule. The exception is, however, only apparent ; the twenty perspinal pores of the shell show that they were originally in connection with twenty centrogenous acanthin spines, and that these have disappeared by retrograde metamorphosis.
107. Perigenous and Centrogenous Skeletons.-Much more important than the topographical relation of the skeleton to the central capsule, according to which the Ectolithia and Entolithia are separated from each other (§ 106), is the original development of the skeleton within or without the central capsule, which gives rise to the distinction between perigenous and centrogenous skeletons. Centrogenous skeletons are found only in the ACANTHARIA, which are further distinguished from all other Radiolaria by their skeleton being formed of acanthin ; in all ACANTHARIA the formation of the skeleton begins in the middle of the central capsule, from which twenty (the number is inconstant only in the small group Actinelida) radial spines are centrifugally developed. The three other legions, on the contrary, possess on the whole a perigenous skeleton, which originally develops outside the central capsule and never in its middle. In the NASSELLARIA and PHAEODARIA the skeleton retains this extracapsular position, as also in the Beloidea and part of the Sphaerellaria among the SPUMELLARIA; in the great majority of the latter, however, the primary perigenous skeleton is subsequently enveloped by the growing central capsule, so that it lies partially within it (§ 109).
108. Polyphyletic Origin of the Skeleton.-The skeleton of the Radiolaria has undoubtedly originated polyphyletically, for it is impossible to derive its manifold varieties from a single ground-form, or to regard them as modifications of one type. It is much more probable that the different skeletonless Radiolaria have entered upon different ways of skeleton formation quite independently of each other. At the outset it is quite clear that the skeletons of the four legions have originated independently of each other. Further, it is certain that within the legion of the SPUMELLARIA the Beloid skeletons of the Collodaria are not connected with the Sphaeroid skeletons of the Sphaerellaria and the forms derived from them (see § 109). In the same way the skeletons of the PHAEODARIA are polyphyletic ; probably in this legion the Beloid, Sphaeroid, Cyrtoid, and Conchoid skeletons have been developed quite independently (see § 112). In the NASSELLARIA, on the other hand, it is possible that all the skeletal forms are to be derived monophyletically from a single simple primitive form (either the sagittal ring or basal tripod?) (see § 111). Still more probable is it that the ACANTHARIA have arisen monophyletically, for all the forms of their acanthin skeleton may be derived without violence from Actinelius (see § 110).
109. The Skeleton of the Spumellaria-The skeletons of the SPUMELLARIA or PERIPYLEA consist of silica, and are very different and of independent origin in the two orders of this legion. The first order, Collodaria, have either no skeleton whatever (Colloidea, p. 10, Pls. 1, 3), or their skeleton is Beloid, a loose extracapsular envelope of spicules, consisting of numerous unconnected portions ; the separate parts are usually disposed tangentially, either as simple or compound siliceous spicules (Beloidea, p. 28, Pls. 2, 4). The second order of SPUMELLARIA, on the other hand (Sphaerellaria, p. 49), develops a siliceous lattice-shell, which consists of a single piece, and is remarkable for the extraordinary variety of its forms (pp. 50-715, Pls. 5-50). To this order belong not less than three hundred genera and seventeen hundred species of the Challenger Radiolaria (that is, about two-fifths of all the genera and species). In spite of this extreme richness in different forms, this large group must be regarded as monophyletic, since all its forms may be quite naturally derived from a common stem-form, a simple lattice-sphere (Cenosphaera, p. 61, Pl. 2). The twenty-eight families of Sphaerellaria may be distributed in four suborders, among which the Sphaeroidea constitute the stem-forms, since they retain the original spherical shape (Pls. 5-8, 11-30). In the other three suborders a vertical main axis is developed, which in Prunoidea is longer, in Discoidea shorter than the other axes of the shell. Hence the shell of the Prunoidea (p. 284, Pls. 13, bis, 17, 39, 40) is ellipsoidal or cylindrical, that of the Discoidea, on the other hand, lenticular or discoidal (p. 402, Pls. 31-38, 41-48). Finally, the shell of the fourth suborder, Larcoidea, is lentelliptical ; it has the ground-form of a triaxial ellipsoid, and is characterised by the possession of three unequal dimensive axes, or three isopolar axes of different lengths perpendicular to each other (p. 599, Pls. 9, 10, 49, 50).
110. The Skeleton of the Acantharia.-The skeletons of the ACANTHARIA or ACTIPYLEA are distinguished from those of all other Radiolaria by two very important peculiarities ; in the first place, they consist not of silica but of a peculiar organic substance, Acanthin, and secondly, their development is centrogenous, numerous radial spines or acanthin spicules being formed which are united in the middle of the central capsule. Hence the ACANTHARIA are the only Radiolaria in which the skeleton originates from the first in the middle of the central capsule. The number of radial spines is primitively indefinite, variable, and often considerable (more than a hundred), but in the great majority it is limited to twenty. In accordance with this the legion may be divided into two orders, the more archaic small group Adelacantha, with an indefinite number of spines, and the more recent group, Icosacantha, which has been developed from them and possesses twenty regularly disposed spines ; of the three hundred and seventy-two species of ACANTHARIA which have been hitherto described, about five per cent. belong to the former, about ninety-five per cent. to the latter division (see note A, below). The numerous genera of Icosacantha may then be again divided into two suborders, of which the Acanthonida (p. 740, Pls. 130, 131, 132) produce no complete lattice-shell, and thus agree with the Actinelida, with which they maybe united as Acanthometra in the broader sense (or ACANTHARIA without a lattice-shell). The Acanthophracta, on the other hand (p. 791, Pls. 133 -140), produce a complete lattice-shell, usually by means of two opposite or four crossed transverse processes, which arise from each radial spine and unite with each other (see note B, below). In most Acanthophracta the lattice-shell remains single ; only in the Phractopeltida does it consist of two concentric lattice-spheres (p. 847, Pl. 133, figs. 1-6). Furthermore, the whole order Acanthophracta may be subdivided into two suborders according to the different ground-form of the lattice shell; this remains spherical in the Sphaerophracta (the three families Sphaerocapsida, Dorataspida, Phractopeltida, Pls. 133-138). On the other hand, it assumes another form in the Prunophracta ; it becomes ellipsoidal in the Belonaspida (Pl. 136, figs. 6-9), discoidal or lentiform in the Hexalaspida (Pl. 139); and finally takes the shape of a double cone in the Diploconida (Pl. 140).
A. The group Adelacantha consists only of the suborder Actinelida, with the three families Astrolophida, Litholophida, and Chiastolida (p. 728, Pl. 129, figs. 1-3) ; tbe number of the radial spines is very different and variable, sometimes only from ten to sixteen, but usually from thirty to fifty, and often more than one hundred ; they are generally irregularly distributed, and not as in the second main division. This latter, the Icosacantha, always possesses twenty radial spines, which are regularly disposed according to a constant law, the so-called "Müllerian" or "Icosacanthan" law ; the twenty spines are always so placed between the poles of a spineless axis that they form five zones each of four spines ; the four spines of each zone are equidistant from each other, and also from the same pole, and alternate with those of the neighbouring zones, so that the whole twenty lie in four meridian planes, which cut out an angle of 45° (compare pp. 717-722, Pls. 130-140). In spite of the manifold variations in form which are developed in the Icosacantha, they may all be derived from a common stem-form, Acanthometron (p. 742), since the law of distribution of the twenty spines is constantly inherited.
B. An exception is found in the peculiar family Sphaerocapsida (p. 797,
Pl. 133, figs. 7-11 ; Pl. 135,
figs. 6-10). Here the shell is composed of innumerable small, perforated
plates, which arise on the surface of the calymma independently of the
spines.
111. The Skeleton of the Nassellaria.-The skeletons of the NASSELLARIA or MONOPYLEA consist of silica, and are never composed of separate portions, but constitute always a single continuous piece. The ground-form is originally monaxon, corresponding to that of the central capsule, with a constant difference between the two poles of the vertical main axis. The ground-form is never spherical or polyaxon as in the lattice-shells of the SPUMELLARIA, and the skeleton never consists of hollow tubes as in the PHAEODARIA. The legion NASSELLARIA may be divided into two orders ; in the Plectellaria (three suborders Nassoidea, Plectoidea, Stephoidea) the skeleton does not form a complete lattice-shell ; in the Cyrtellaria, on the other hand, which are derived from these, the siliceous skeleton forms a complete lattice-shell enclosing the central capsule. The number of forms thus developed is astonishingly great, so that among the NASSELLARIA no less than two hundred and seventy-four genera and sixteen hundred and eighty-seven species may be distinguished, almost as many as in the Sphaerellaria. In spite of this great variety of forms the legion MONOPYLEA is probably monophyletic ; at least all the different skeletal forms may be derived from three elements which are combined in the most manifold fashion ; (1) the sagittal ring, a simple siliceous ring, which lies vertically in the sagittal plane of the body, encircles the central capsule and comes into contact with it at the basal pole of the main axis (§ 124); (2) the basal or oral tripod, composed of three diverging radial spines, which meet in the middle of the basal pole of the central capsule (or in the centre of the porochora) (§ 125) ; (3) the cephalis, or lattice-head, a simple ovoid or subspherical lattice-shell, which encloses the central capsule and stands in connection with it at the basal pole of its main axis. Any one of these three important structural elements of the NASSELLARIAN skeleton may possibly be the starting-point for all the remaining forms of the MONOPYLEA ; the great difficulty in their phylogenetic derivation lies in the facts that, on the one hand, any one of the three elements may alone constitute the skeleton, and on the other hand, in the great majority of the legion, two or three are united together (compare §§ 182-185).
112. The Skeleton of the Phaeodaria.-The skeleton of the PHAEODARIA or CANNOPYLEA is always extracapsular, usually consists of a silicate of carbon (more rarely of pure silica), and in the majority of the legion is composed of hollow cylindrical tubes, whose siliceous wall is very thin, and whose lumen is filled with gelatinous material (§ 127). The manifold and remarkable skeletal forms occurring in this legion are not monophyletic, since they cannot be derived from a common stem-form ; they are, on the contrary, polyphyletic, various skeletonless PHAEODARIA (Phoeodinida) have independently acquired skeletons of different form and composition. The legion PHAEODARIA can be subdivided into four orders, the skeletons of which present the following important distinctions :- (1) The Phaeocystina possess only incomplete Beloid skeletons (§ 115), composed of many separate pieces, sometimes tangentially (Cannorhaphida, Pl. 101), sometimes radially arranged (Aulacanthida, Pls. 102-105). (2) The Phaeosphaeria form Sphaeroid skeletons (§ 116), usually only a simple lattice-shell without special aperture (Pls. 106-111); two concentric shells united by radial bars occur only in the Cannosphaerida (Pl. 112). (3) The Phaeogromia are distinguished by the formation of a simple Cyrtoid skeleton (§ 123) resembling that of the Monocyrtida ; the monothalamus lattice-shell is usually ovoid or helmet-shaped, more rarely polyhedral or almost spherical ; a vertical main axis can always be distinguished, at the basal pole of which is an aperture usually armed with teeth or spines (Pls. 99, 100, 113-120). (4) The Phaeoconchia are distinguished from all other Radiolaria by the possession of a bivalved shell like that of the Conchifera ; the two valves of this Conchoid skeleton must be distinguished as dorsal and ventral, as in the Brachiopoda (Pls. 121 -128). The fifteen families of PHAEODARIA which are arranged in the four orders just mentioned, present such great differences among themselves, that the skeleton must be regarded as probably polyphyletic even within the limits of each order.
113. Types of Skeletal Formation.-No less than twelve different principal forms may be distinguished as morphological types of the formation of the skeleton in the Radiolaria ; some of these are peculiar to a single legion or even to a smaller group ; but sometimes the same form occurs in several legions. Some types occur only in an isolated manner, independently of the others, but most exist in various combinations with other types. Of the twelve described below the Conchoid and Cannoid occur only in the PHAEODARIA ; the Plectoid and Circoid only in the NASSELLARIA ; the Astroid only in the ACANTHARIA ; the remaining seven types are found in several legions in the same form and hence are polyphyletic.
114. The Astroid Skeleton.-Under the name "Astroid" we place the peculiar star shaped skeletons of the ACANTHARIA in opposition to those of all other Radiolaria, for they are separated from them not only fundamentally by reason of the chemical nature of their substance (Acanthin, § 102), but also by their centrogenous origin, and the resulting stellate form (Pls. 129-140). The ACANTHARIA are the only Radiolaria in which the skeleton arises within the central capsule by the formation of numerous rays or radial spines of acanthin which project on all sides from the centre. Originally these are united at this point, their conical or pyramidal points meeting and being supported one upon another. In the great majority of ACANTHARIA this loose apposition is constant, so that when the soft parts are destroyed the skeleton falls to pieces. Only in a few forms in this legion are the central ends of the spines fused so that the whole skeleton forms a connected star (Astrolithium). The small group Chiastolida (or Acanthochiasmida) is characterised by the fact that the two rays which are opposite to one another in each axis unite and form a diametral bar. The skeleton is almost always composed of twenty radial spines, which are regularly disposed (Icosacantha), only in the small primitive group Actinelida is the number variable (Adelacantha, § 110).
115. The Beloid Skeleton.-As Beloid or spicular skeletons are grouped together all those which consist of several disconnected portions ; these always lie outside the central capsule, either within the calymma or on its surface. Such extracapsular Beloid skeletons are entirely wanting in the ACANTHARIA and NASSELLARIA ; they occur only in the Beloidea among the SPUMELLARIA, and in the Phaeocystina among the PHAEODARIA ; the individual Beloid portions of the former are solid, those of the latter hollow. In both groups the simplest forms of the separate portions are simple unbranched needles (Thalassosphaera, Thalassoplancta, Physematium, Belonozoum, among the SPUMELLARIA; Cannobelos and Cannorrhaphis among the PHAEODARIA) ; usually these spicules are disposed tangentially over the surface of the calymma. Among the Beloidea branched spicules occur more commonly than these simple ones ; they are either stellate (with many rays united in a centre) or twin-like, with a tangential bar, from each pole of which two or three (seldom more) radial branches project (Pls. 2, 3, 4), Among the PHAEODARIA the subfamily Dictyochida is characterised by the annular shape of its Beloid portions, either simple rings, or hat-shaped or pyramidal bodies with a latticed cap over the ring (Pl. 101, figs. 3-14 ; Pl. 114, figs. 7-13). The family Aulacanthida among the PHAEODARIA, alone possesses hollow radial tubes, which penetrate the whole calymma, and project distally over its surface, whilst their proximal ends rest upon the surface of the central capsule. Although in these cases the enclosed proximal end is always simple, the free distal end develops the most various processes in adaptation to its prehensile functions (Pls. 102-105).
116. The Sphaeroid Skeletons or Lattice-Spheres-The "lattice-spheres" or sphaeroid skeletons are the simplest and most primitive forms of lattice-shells, and are widely dis tributed in the three legions SPUMELLARIA, ACANTHARIA, and PHAEODARIA, whilst they are entirely wanting in the NASSELLARIA. The round lattice-shell is either a true sphere in the geometrical sense, or an endospherical polyhedron, i.e., a polyhedron, all whose angles lie in the surface of a sphere (§ 25). In general, primary and secondary lattice spheres may be distinguished, of which the former are secreted on the outer surface of the primary, the latter on that of the secondary calymma (§ 85). Furthermore, simple and compound lattice-spheres may be distinguished, the latter of which consist of two or more concentric lattice-spheres firmly united by radial bars ; in such cases the inner most lattice-sphere is always to be regarded as the oldest or primary, all the succeeding ones as secondary, and the outermost as the youngest (§ 129). The simple lattice-spheres are usually to be regarded as primary ; they may, however, occasionally be secondary, in which case the primary shell, originally enclosed, has been lost by degeneration (as, for example, in the case of the Aulosphaerida and some Sphaerellaria).
117. The Lattice-Spheres of the Spumellaria.-The lattice-spheres or Sphaeroid skeletons of the SPUMELLARIA exhibit in spite of their simple type of structure, an extra ordinary variety in the formation of the lattice-work and radial apophyses, so that in the systematic portion of this work no less than one hundred and seven genera and six hundred and fifty species are distinguished ; these are united in one suborder, the Sphaeroidea (pp. 50-284, Pls. 5-8, 11-30). It may be divided into two main divisions, the Monosphaerida with a single primary lattice-sphere (Pls. 12-14, 21, 26, 27), and Pliosphaerida (or Sphaeroidea concentrica) whose skeleton consists of two or more concentric lattice-spheres united by radial bars. The latter are subdivided into Dyosphaerida with two concentric lattice-spheres (Pls. 16, 19, 20, 22, 28); Triosphaerida, with three spheres (Pls. 17, 24, 29); Tetrasphaerida, with four (Pls. 23, 30); Polysphaerida, with five or more (Pls. 15, 23); and Spongosphaerida, with spongy lattice-spheres (Pls. 18, 25). A special group is made up of the simple lattice-spheres of the social Collosphaerida (or Sphaeroidea polyzoa) (Pls. 5-8); these are usually more or less irregular, and characterised by the development of peculiar tubular processes ; the latter are generally wanting in the Sphaeroidea monozoa, whose lattice-shell is very regularly formed. This distinction is interesting and important, inasmuch as the regular lattice-spheres are explained by the independent development of the free-swimming Monozoa, whilst the irregular spheres are due to the mutual dependence of the social Polyzoa.
118. The Lattice-Spheres of the Acantkaria.-The lattice-shells or Sphaeroid skeletons of the ACANTHARIA are immediately distinguishable from those of all other Radiolaria by their centrogenous development and the central union of the radial spines by which they are supported ; the only exception is furnished by the remarkable genus Cenocapsa (Pl. 133, fig. 11), in which the radial spines are absent, not primitively, however, but in consequence of degeneration ; for the twenty cross-shaped perspinal pores, originally due to the twenty radial spines, are still present. In the most nearly allied genera, Porocapsa (Pl. 133, fig. 7) and Cannocapsa (Pl. 133, fig. 8), the proximal part of the twenty radial spines is still present, while their distal portion has degenerated ; hence in this case they do not stand in direct communication with the spherical shell. On the other hand, this primitive connection persists in the genera Astrocapsa (Pl. 133, figs. 9,10), and Sphaerocapsa (Pl. 135, figs. 6-10). The five genera just mentioned form the peculiar family Sphaerocapsida (pp. 795-802); the spherical shell is in these cases composed of very numerous small plates disposed like a pavement, each plate or aglet being perforated by a pore canal ; in addition to which there are twenty larger (perspinal) pores (or twenty cross-shaped groups each of four aspinal pores) at those important points where primitively the twenty radial spines penetrate the calymma. This peculiar porous "pavement shell" has probably been developed (independently of the twenty radial spines) upon the calymma of the Acanthonida (Acanthonia, p. 749) by the action of the sarcodictyum ; it has, therefore, quite a different morphological significance from the spherical lattice-shell of the Dorataspida, which is composed of tangential apophyses of the twenty Acanthonid spines (pp. 802-847, Pls. 134-138). Each radial spine here forms either two opposite or four crossed transverse processes, and since their branches spread over the surface of the spherical calymma and are united suturally at their extremities, the peculiar lattice-sphere of the Dorataspida arises. This extensive family is again divided into two subfamilies :-the Diporaspida (Pls. 137, 138) possess always only two opposite apophyses, and form by the union of their branches two opposite primary apertures or aspinal meshes. The Tessaraspida, on the other hand (Pls. 135, 138), have always four crossed transverse processes, and form by their union four primary aspinal meshes. From the Diporaspida are probably to be derived the Phractopeltida (p. 847, Pl. 133, figs. 1-6), the only ACANTHARIA which possess a double lattice-sphere ; their double concentric spherical shell may be compared with that of the Dyosphaerida.
119. The Lattice-Spheres of the Phaeodaria.-The lattice-spheres or Sphaeroid skeletons of the PHAEODARIA, which are generally developed quite regularly, though occasionally in a modified form, fall in the order Phaeosphaeria into two groups of very different structure, each of which includes two families. The first group (Phaeosphaeria inarticulata) contains the families Orosphaerida (Pls. 106, 107) and Sagosphaerida (Pl. 108); the lattice-work of the former consists of irregular polygonal meshes and very coarse, partially hollow trabeculae ; in the latter, on the other hand, it consists of triangular meshes and very slender filiform trabeculae ; in both families the whole sphaeroid skeleton forms a single unsegmented piece as in most Sphaeroidea. In the second group of Phaeosphaeria (Phaeosphaeria articulata), on the other hand, the lattice-sphere is segmented in quite a peculiar manner, and composed of hollow cylindrical tangential tubes, which are separated by astral septa at the nodal points of the network ; this remarkable structure characterises the two families, Aulosphaerida (Pls. 109, 110, 111) and Cannosphaerida (Pl. 112); the segmented lattice-sphere of the former is simple and hollow ; while that of the latter is connected by centripetal radial tubes with a simple concentric inner shell, which is sometimes solid, sometimes latticed, and provided with a main opening corresponding to the astropyle of the enclosed central capsule. Since in the Aulosphaerida also, hollow centripetal radial tubes project from the segmented lattice sphere, it is possible that they have been derived from the Cannosphaerida by the loss of the primitive internal shell. A special peculiarity of many Phaeosphaeria (Oroscena, Sagoscena, Auloscena, &c.) consists in the fact that the whole surface of the lattice sphere is regularly covered with pyramidal or tent-shaped prominences (Pl. 106, fig. 4 ; Pl. 108, fig. 1; Pl. 110, fig. 1). A simple lattice-sphere quite similar to that of most Monosphaerida also constitutes the skeleton of the Castanellida (Pl. 113), but since it possesses a special main-opening, it must be referred promorphologically to the Cyrtoid shells of the Phaeogromia.
120. The Prunoid Skeleton or Lattice-Ellipsoid.-The "lattice-ellipsoids" or Prunoid skeletons have arisen from the lattice-spheres or Sphaeroid skeletons by more growth and elongation of one axis ; this is the main axis of the body and is probably always vertical ; its two poles are commonly equal. The Prunoid skeleton is either a true ellipsoid in the geometrical sense or an "endellipsoidal polyhedron" (i.e., a polyhedron, all the angles of which lie in an ellipsoidal surface). By further elongation of the main axis, the ellipsoidal form passes over into the cylindrical, the polar surfaces of the cylinder being usually rounded, rarely truncated. The rich order Prunoidea (pp. 284-402) contains numerous modifications of this form of shell which arise on the one hand by the formation of transverse constrictions, on the other by the apposition of concentric secondary shells. In respect of the latter, simple and compound Prunoid shells can be distinguished as in the case of the Sphaeroid shells. In the compound Prunoid shells either all the concentric lattice-shells may be ellipsoidal or the inner may be spherical. More important differences are found in the transverse annular constric tions, which give the Prunoid skeleton a segmented appearance ; in this respect, three principal forms may be distinguished (p. 288) :-(A) Monoprunida, with unsegmented shell, having no transverse constriction (Pls. 15, 16, 17); (B) Dyoprunida, having a shell with two segments and one (equatorial) transverse constriction (Pl. 39); (C) Polyprunida, with three or more parallel transverse constrictions, by means of which the shell is divided into four or more segments (Pl. 40). In the same manner as the Prunoidea have arisen from the Sphaeroidea among the SPUMELLARIA by greater development of the vertical main axis, the ellipsoidal Belonaspida have arisen from the spherical Dorataspida among the ACANTHARIA (p. 859 ; Pl. 136, figs. 6-9 ; Pl. 139, figs. 8, 9). The main axis of the ellipsoid in this case is always occupied by the opposite equatorial spines of the hydrotomical axis (pp. 719, 860). In the legion PHAEODARIA a similar prolongation of the main axis rarely occurs ; it is found, however, in Aulatractus (Pl. 111, figs. 6, 7), the lattice-shell of this Aulosphaerid being sometimes truly fusiform, sometimes rather ellipsoidal or even double-conical.
121. The Discoid Skeletons or Lattice-Discs.-The "lattice-discs" or Discoid skeletons are characteristic of the SPUMELLARIAN group Discoidea, and have arisen from the lattice-spheres of the Sphaeroidea by a less development of one axis, which is the main axis of the body, and is probably usually vertical ; its two poles are always equal. The Discoid lattice-shell is either a biconvex lens (with a thin margin), or a plane disc (a shortened cylinder with thick margin), or some form intermediate between the two. All Discoid shells show a horizontal median plane or equatorial plane, by which they are divided into two equal halves, an upper and lower ; the margin of the lens itself is originally the equator. The main axis, the shortest of all the axes of the shell, stands vertically in the centre of the equatorial plane. Among the PHAEODARIA Discoid shells rarely occur (Aulophacus), as also among the ACANTHARIA (Hexalaspida).
122. The Larcoid Skeleton or Lentelliptical Lattice-Shell.-The lentelliptical lattice-shells, which may be shortly designated "Larcoid," are especially characteristic of the Larcoidea, a large order of SPUMELLARIA (pp. 599-715; Pls. 9, 10, 49, 50). In addition they recur among the ACANTHARIA, in the small family Hexalaspida (p. 872, Pl. 139), and the family Diploconida (p. 881, Pl. 140), which is derived from it. These lentelliptical lattice-shells are all characterised by the clear differentiation of three unequal, but isopolar dimensive axes, i.e., the three geometrical axes, perpendicular to one another, which determine the form of the shell, are of unequal length ; the two poles of each are, however, equal. The geometrical ground-form is, therefore, a triaxial ellipsoid (§ 34). In the rich order Larcoidea the lentelliptical lattice-shell shows many variations in its development.
123. The Cyrtoid Skeleton.- Cyrtoid skeletons are those lattice-shells which possess a vertical main axis with two different poles (Monaxonia allopola) ; the upper pole is usually termed the apical, the lower the basal. Such Cyrtoid shells are characteristic of the great majority of the NASSELLARIA or MONOPYLEA (and especially of the Cyrtellaria) ; they are also found in a large division of the PHAEODARIA (the Phaeogromia), and in some SPUMELLARIA. In general the manifold Cyrtoid shells may be divided into two large groups, those with one and those with several chambers. The monothalamous Cyrtoid shells are usually ovoid, conical, cap- or helmet-shaped ; their internal cavity is simple, without constrictions or septa. Among the NASSELLARIA they occur in the Monocyrtida (Pls. 51-54, 98), where they have received the name "Cephalis." A form of shell, essentially the same, is found amongst the PHAEODARIA in the order Phaeogromia, more especially in the Challengerida (Pl. 99), Medusettida (Pls. 118-120), and Tuscarorida (Pl. 100), many of these latter closely resembling many Monocyrtida. Such monothalamous Cyrtoid shells occur much more rarely among the SPUMELLARIA (e.g., among the Prunoidea in Lithapium, Lithomespilus, Druppatractus, Pls. 13, 14, &c.). Polythalamous Cyrtoid shells (Pls. 55-80) occur exclusively in the NASSELLARIA, and exhibit in this legion an astonishing variety of structure ; they are distinguished from the monothalamous forms by the development of internal septa, or of annular incomplete diaphragms, which usually correspond to the external constrictions ; their interior is thus divided into two or more communicating compartments. Among the polythalamous Cyrtoid shells may be distinguished three principal groups, the Stichocyrtid, Zygocyrtid, and Polycyrtid. Zygocyrtid shells are characteristic of the Spyroidea (Pls. 84-90), and are distinguished by a bilobate cephalis (cephalis bilocularis) ; the median sagittal ring, or a corresponding constriction, divides the shell into right and left compartments. Polycyrtid shells (Pl. 96) are peculiar to the Botryodea, and characterised by a multilobate cephalis (cephalis multilocularis). Stichocyrtid shells are those in which the primary cephalis remains simple, and new joints are successively added to its basal pole ; such shells occur in the majority of the Cyrtoidea. Secondary chambers are sometimes added in the other two groups (Botryodea and Spyroidea). When, as often happens in these polythalamous Cyrtoid shells, two or three distinct joints follow each other, the first is called the "cephalis," the second the "thorax," and the third the "abdomen" (Tricyrtida, Pls. 64-75).
124. The Circoid Skeleton.-This is a very important and remarkable type of skeletal formation, which occurs exclusively in the legion NASSELLARIA, where it plays a very prominent part ; its characteristic element is the "sagittal ring," a simple, vertical, siliceous ring, which surrounds the central capsule in its sagittal plane, and is specially differentiated in its basal portion. This "primary sagittal ring" whose vertical allopolar main axis coincides with that of the Monopylean central capsule embraced by it, is characteristic of all members of the order Stephoidea (p. 931, Pls. 81-83, 92-94); here it forms by itself the skeleton of the Stephanida (Pl. 81); in the Semantida (Pl. 92) it is combined with a horizontal basal ring, in the Coronida (Pls. 82, 93) with a vertical frontal ring, and in the Tympanida (Pls. 83, 94) with two horizontal rings, an upper mitral and a lower basal. In the great majority of these Stephoidea there often develop in definite places characteristic processes or apophyses, whose branches combine to form a loose tissue or an incomplete lattice-shell. This becomes complete in the Cyrtellaria, the majority of which retain more or less distinct traces of the sagittal ring. Hence the skeletons of all NASSELLARIA may be derived monophyletically (Hypothesis A, p. 893) from a simple sagittal ring (Archicircus and Lithocircus, Pl. 81). This theory, however, encounters the great difficulty that in many Stephoidea (Cortina, Cortiniscus, &c.) it is combined in a remarkable manner with the basal tripod of the Plectoidea whilst in these latter it is entirely wanting (compare p. 894).
125. The Plectoid Skeleton.-Those forms are distinguished as Plectoid in which three, four, or more radial siliceous spines proceed from a common point, which lies excentrically outside the central capsule and at the basal pole of its vertical allopolar main axis. This peculiar type of skeletal formation only occurs in the legion NASSELLARIA, and is specially characteristic of the order Plectoidea (p. 898, Pl. 91). But since the essential elements of this remarkable skeleton also occur in many other NASSELLARIA, sometimes combined with the Circoid, sometimes with the Cyrtoid skeleton, it perhaps has a fundamental significance in this legion ; at all events it is possible to derive monophyletically all the other forms of this legion from it. (Hypothesis B, p. 893). The simplest form of the Plectoid skeleton is a tripod, the three feet of which either lie in a horizontal plane (Triplagia, Pl. 91, fig. 2), or correspond to the three edges of a low pyramid (Plagiacantha). A fourth ray is sometimes added, which stands vertically upon the summit of the pyramid (Plagoniscus, Plagiocarpa, Pl. 91, figs. 4, 5). In other Plectoidea three secondary rays are intercalated between the three primary (Hexaplagida, &c.) ; seldom the number is greatly increased (Polyplagida, &c.). The rays are rarely simple, but usually branched ; in the Plagonida (Pl. 91, figs. 2-6) the branches remain free ; in the Plectanida (Pl. 91, figs. 7-13) they are united to form a loose wicker-work. From such a web a perfect Cyrtoid shell may arise. Several forms of Plagonida may also be readily confounded with the isolated triradiate or quadriradiate spicula of many Beloid skeletons (Sphaerozoum, Lampoxanthium, &c.).
126. The Spongoid Skeleton.-From the simple lattice-skeleton
which the majority of Radiolaria possess, some of them develop a spongy
shell ; the trabeculae of the lattice-work, situated in one plane in the
former, are developed in the latter in different planes and cross irregularly
in all directions ; thus arises a kind of wicker-work of more or less spongy
structure, usually with very thin trabeculae and irregular meshes. Such
Spongoid shells are most common among the SPUMELLARIA, especially in the
Sphaeroidea (Spongosphaerida, Pl. 18)
and Discoidea (Spongodiscida, Pls. 41-47),
more rarely in the Prunoidea and Larcoidea. Lattice-work of similar spongy
structure occurs very seldom among the NASSELLARIA, e.g.,
in some
Plectoidea (Pl. 91) and Cyrtoidea
(Spongocyrtis,
Spongopyramis, Spongomelissa,
&c., Pl. 56,
fig. 10 ; Pl. 64, figs. 5-10,
&c.). Among the PHAEODARIA spongy skeletons are very rare ; they are
to be seen in some Phaeosphaeria (Oroplegma, Pl. 107,
fig. 1; Sagoplegma,Pl. 108,
fig. 2 ; Auloplegma, Pl. 111,
fig. 8). No Spongoid skeletons are known among the ACANTHARIA.
127. The Cannoid Skeleton.-Cannoid or tubular skeletons are those which are composed of hollow tubes ; they occur exclusively in the PHAEODARIA or CANNOPYLEA. Tubular processes, nevertheless, occur in some other Radiolaria, as, for example, among the SPUMELLARIA in a portion of the Collosphaerida (Siphonosphaera, Caminosphaera, Pls. 6, 7), and of the Prunoidea (Pipetta, Cannartus, &c., Pl. 39, figs. 6-10, &c.), also among the NASSELLARIA in Theosyringium (Pl. 68, figs. 4-6), Cannobotrys (Pl. 96, figs. 3, 4, 8-11, 20-22), &c. In all these cases, however, the tubes are direct processes of the cavity of the shell, the trabeculae of the lattice-work being solid. Only in the CANNOPYLEA are the lattice-bars themselves, the radial spines and appendicular organs, generally tubular (hence the designation "Pansolenia"). The lumen of the thin-walled siliceous tubes is filled with jelly, and hence the specific gravity of the relatively large skeleton is considerably diminished. This peculiarity is not found in all CANNOPYLEA ; it is wanting in all Sagosphaerida and Concharida, as well as in a part of the Orosphaerida and Castanellida ; in the latter there are found intermediate stages between hollow and solid skeletal rods. Very often a fine siliceous thread runs in the axis of the tubes, which is connected with its wall by lateral branches (Pl. 110, figs. 4, 6 ; Pl. 115, figs. 6, 7). More seldom the tubes are divided by horizontal septa into a series of chambers (Medusettida, Pls. 118-120). The two families Aulosphaerida (Pls. 109-111) and Cannosphaerida (Pl. 112) are distinguished from all other PHAEODARIA by the fact that their tubes are separated by astral septa in the nodal points of the lattice-shell (§§ 112, 134).
128. The Conchoid Skeleton.-By the name "Conchoid skeletons" are distinguished the bivalved lattice-shells which occur exclusively in the legion PHAEODARIA ; they are quite characteristic of the Phaeoconchia or Phaeodaria bivalvia, which embrace three families :-Concharida (Pls. 123, 124, 125), Coelodendrida (Pls. 121, 122), and Coelographida (Pls. 126, 127, 128). The two valves of the lattice-shell of the Concharida are simple, hemispherical, or boat-shaped, whilst in the Coelodendrida and Coelographida tubes grow out from them, which branch and usually give rise by anastomosis to a second external bivalved shell. In all Phaeoconchia the two valves are so disposed about the central capsule that an open slit remains between them, into which open the apertures of the central capsule ; and since all these Phaeodaria conchoidea are TRIPYLEA, with three typical openings in the central capsule, and since the two lateral accessory openings lie at either side of the aboral pole, and the unpaired main-opening at the oral pole of the main axis, it follows that the two valves are to be regarded as dorsal and ventral as in the Brachiopoda (not right and left as in the Lamellibranchiata). The dorsal and ventral valves are usually equal, but in a portion of the Concharida they present constant differ ences. In this family the two valves are attached to each other by their free edges, just as in the bivalved Mollusca and Diatoms ; and these edges may either be smooth (Conchasmida, Pl. 123, figs. 1-6), or dentate (Conchopsida, Pls. 124, 125); the valvular connection of the latter is sometimes strengthened by a special ligament which unites the two valves at the aboral pole (Pl. 123, figs. 8, 9). The form of the valve is sometimes hemispherical, sometimes boat-shaped, with a sagittal keel.
129. Medullary and Cortical Shells.-In all Radiolaria whose skeleton consists of a double shell or of two concentric lattice-shells united by radial bars, an inner medullary shell (testa medullaris) and an outer cortical shell (testa corticalis) may be distinguished (see note A, below). The medullary shell is usually to be regarded as a primary, the cortical as a secondary structure. Such double shells occur among the SPUMELLARIA in the Dyosphaerida (Pls. 19,20), as well as in many Prunoidea (Pls. 39, 40), Discoidea (Pls. 33, 34), and Larcoidea (Pls. 9, 10); among the ACANTHARIA only in the family Phractopeltida (Pl. 133); among the NASSELLARIA only in very few Cyrtoidea (e.g., Periarachnium, Pl. 55, fig. 11), and finally among the PHAEODARIA in the Cannosphaerida (Pl. 112) as well as in part of the Coelodendrida (Pl. 121) and Coelographida (Pls. 127, 128). In most cases (if not always?) the cortical shell arises by the growth of radial spines from the surface of the medullary shell ; these become united at equal distances from the centre by transverse apophyses, the surface of the secondary calymma furnishing the basis for their secretion (§ 85). Nevertheless, it seems that in many Sphaerellaria the formation of the whole cortical shell proceeds simultaneously (at a definite dictyotic period) like that of the primary medullary shell (see note B). Whilst in the PHAEODARIA, ACANTHARIA, and NASSELLARIA, at most two concentric shells are formed, in many SPUMELLARIA their number increases continuously with additional growth ; in many Sphaerellaria it rises to four, eight, or even more, as well as in many Discoidea (if the concentric, peripherally disposed rings of chambers be regarded as incomplete flattened shells). In these cases either only the innermost primary lattice-shell is to be styled " medullary shell," or at most the two innermost (inner and outer medullary shells), all the others being cortical.
A. The distinction between medullary and cortical shells was originally based in my Monograph (1862, p. 50) upon the topographical relation of the lattice-shells to the central capsule, inasmuch as I regarded all intracapsular shells as medullary, all extracapsular as cortical. Hertwig, however (1879, p. 122), rightly pointed out that this distinction is unpractical, "because the same lattice shell in the same species may lie within or without the central capsule, according to the size of the latter." He proposes, therefore, to restrict the term medullary shell to the innermost, and to call all the others cortical ; a course which seems justified by the special significance of the primary innermost lattice-shell ("as the point of origin of the radial spines "). But in most Sphaerellaria which form three or more concentric shells, the two innermost, which lie near together within the central capsule, are very different in size and dictyosis from all the others which lie outside, and are separated by wider interspaces (compare Pls. 17, 24, 29-32, 40, &c.). In these cases it appears better to regard the two inner as inner and outer medullary shells, and all the others as cortical shells. The character of the dictyosis in the intracapsular and extracapsular shells is often so different that I have made it the basis of separation of Thecosphaera and Rhodosphaera among the Liosphaerida (p. 60), of Elatommatida and Diplosphaerida among the Astrosphaerida (p. 208), &c.
B. R. Hertwig (1879, L. N. 33, pp. 40, 123) separates the true (simultaneously formed) "cortical shells" (e.g., of Actinomma, Cromyomma) from the arachnoid "siliceous networks" (e.g., of Diplosphaera and Arachnosphaera) which are formed by the successive union of tangential apophyses of the radial spines. Whether this principle is right in theory or not, it cannot be carried out practically. Compare also Pl. 25, fig. 4.
130. Dictyosis or Lattice Formation of the Skeleton. In the great majority of Radiolaria the dictyosis or formation of lattice-work, and especially the formation of a variously-shaped "lattice-shell," plays such an important part that the whole class has long been popularly known in Germany by the name "lattice animalcules" (" Gitterthierchen" or "Gitterlinge") (Protista dictyota). The old name Polycystina also (1838), although referring only to the SPUMELLARIA and NASSELLARIA, is derived from the lattice-work of the siliceous skeleton. The extremely various forms in which this is manifested furnish the means of distinguishing species. The specific conformation of the skeletal lattice-work is usually caused by the special disposition of the sarcodictyum (§ 94), whose exoplasmatic threads become silicified or (in the ACANTHARIA) converted into bars of acanthin. In many cases, however, the form of the lattice is mainly dependent upon the situation and form of the radial spines or of special processes from them. With respect to their origin, two varieties of lattice may be distinguished-simultaneous and successive. Simultaneous dictyosis occurs especially in the simple lattice-shells of the Sphaerellaria and PHAEODARIA, where, at a given moment (" dictyotic moment ") the whole lattice of the shell is excreted on the surface of the calymma. Successive dictyosis, on the other hand, is found more particularly in the lattice-shells of the ACANTHARIA (and in the concentric cortical shells of many Sphaerellaria), which develop from separate lattice-plates formed by the apophyses of the radial spines, and hence not at the same moment. The lattice-shells of the Cyrtetellaria, which gradually grow out from a sagittal ring or a basal tripod, arise by successive dictyosis.
131. Dictyosis of the Spumellaria.-Siliceous lattice-structures are wanting in the first section of the SPUMELLARIA, the Collodaria, but in the second section, Sphaerellaria, they are developed in extraordinary variety of details. In spite of this extreme richness in different forms, the lattice-shells of the SPUMELLARIA may all be derived from one and the same primitive ground-form, a simple lattice-sphere with regular hexagonal meshes (Phormosphaera, p. 61, Pl. 12, figs. 9-11; Heliosphoera, Pl. 28, figs. 1-3, &c.). The siliceous bars which bound these regular and subregular meshes are at first exceedingly thin and filiform ; afterwards they become thicker or spread out laterally, whence the meshes often become round with a hexagonal frame (Pl. 12, fig. 5 ; Pl. 28, fig. 1). If the latter vanish, a lattice-shell with simple circular meshes is formed. Very commonly the regular form of the meshes or pores becomes more or less irregular, polygonal, or roundish. Hence, in general, four different principal forms of dictyosis may be distinguished among the SPUMELLARIA ; viz. (1) regular or subregular hexagonal meshes ; (2) regular or subregular circular meshes ; (3) irregular polygonal meshes ; (4) irregular roundish meshes. The three latter forms are to be regarded as secondary, derived from the primary first form. In those SPUMELLARIA which possess several concentric lattice-shells enclosed one within another, either these have all the same form of dictyosis, or the lattice-work of the innermost primary shell is different from that of the outer secondary shells (Pls. 19, 20); sometimes these latter also differ more or less among themselves (§ 129).
132. Dictyosis of the Acanthara.-The lattice-structures of the ACANTHARIA differ essentially from those of other Radiolaria in several particulars. Firstly, they consist not of silica but of acanthin (§ 102); secondly, they are always secondary formations, usually developed from transverse processes of the primary centrogenous radial spines ; thirdly, their formation is not simultaneous (at the same time over the whole shell), but successive (proceeding from the individual radial spines tangentially towards the middle of the intervals) ; fourthly, the configuration of the network is due to the relative position of the spines and the mode of union of their transverse apophyses. Since these are at right angles to the spines, and since the branches of the apophyses are at right angles to them, the original ground-form of their dictyosis is a lattice-work with quadrangular meshes ; these are often quite regular and square (Pl. 130, figs. 5, 6 ; Pl. 136, figs. 2, 9, &c.) ; more commonly they are rectangular or irregularly quadrangular (Pl. 131, fig. 10 ; Pl. 133, figs. 2, 3, &c.). In the majority of the ACANTHARIA the quadrangular form of the meshes passes over into an irregularly polygonal or roundish one (Pls. 137, 138). Very often the primary meshes of the lattice-shells, which immediately surround the radial spines, are larger and more regular (" aspinal pores"), whilst the numerous secondary meshes between them are smaller and irregular (" coronal pores" ; Pl. 135, figs. 1-4 ,&c.).
133. Dictyosis of the Nassellara.-The siliceous lattice-structures of the NASSELLARIA are formed on the whole like those of the SPUMELLARIA, with which they were formerly united under the name "Polycystina." In this group also there may be distinguished as two main forms the regular and the irregular. In the NASSELLARIA the regular lattice- structures generally exhibit hexagonal or circular meshes, whilst the irregular are either polygonal or roundish ; the irregular forms are, however, much more abundant than the regular, and a further distinction from the SPUMELLARIA consists in the fact that the primary skeletal elements, from which the lattice is secondarily developed, exercise a predominant influence upon their form. These primary elements in the majority of the NASSELLARIA are to be seen in two morphobogically most important structures: first, the primary sagittal ring, which embraces the central capsule in the median plane (§ 124); and secondly, the basal tripod (§ 125), whose three diverging rays proceed from the base of the central capsule, whilst commonly a fourth vertical ray supports the dorsal side of latter (compare Pls. 81-91, p. 892). In the majority of the NASSELLARIA these two primary elements appear in combination, whilst in others only one of them is recognisable. In addition there occur numerous monaxon lattice-shells in which neither of these elements can be recognised, but a simple ovoid lattice-shell (cephalis) alone forms the whole skeleton or its primary part (Pl. 51, fig. 13 ; Pl. 98, fig. 13). The great difficulty in the morphological interpretation and phybogenetic derivation of the NASSELLARIAN skeleton lies in the fact that each of these three elements-the primary sagittal ring, the basal tripod, and the latticed cephalis-may form the whole skeleton by itself or be combined with one or both of the others (p. 893). Even nearly related or at all events very similar forms may differ very greatly in this respect. With regard to the manifold forms of their dictyosis it follows that it is partly dependent upon one of the two first elements, partly independent. In the Plectellaria (or those NASSELLARIA which do not possess a complete lattice-shell) the lattice-work is usually irregular and arises by union of the ramifications, which proceed either from the primary sagittal ring (Pls. 81, 82, 92-94) or from the basal tripod (Pl. 91). In the Cyrtellaria (or NASSELLARIA with a complete lattice-shell, Pls. 51-80), on the other hand, the lattice-work is sometimes regular, sometimes irregular, being often very different in the different joints of a segmented shell (Pl. 72); a great part of it arises independently of the two chief morphological elements, and develops according to laws similar to those which regulate the dictyosis of the SPUMELLARIA.
134. Dictyosis of the Phaeodara.-The lattice-structures of the PHAEODARIA, which consist of a silicate of carbon (§ 102), are on the whole not developed in such variety as those of the other Radiolaria, but exhibit several essentially different types of structure, not reducible to a common primitive type of lattice-work. In one portion of this legion there occurs an ordinary simple lattice-work (as in SPUMELLARIA and NASSELLARIA), with solid trabeculae ; of these the Castanellida (Pl. 113) and Concharida (Pls. 123-125) have usually regular or subregular, circular meshes, sometimes hexagonally framed ; the Orosphaerida (Pls. 106, 107) large irregular polygonal meshes with thick trabeculae, the Sagosphaerida (Pl. 108) large triangular meshes with thin filiform trabeculae. The Challengerida (Pl. 99) are characterised by a very delicate regular lattice-work, with minute hexagonal pores, like a Diatomaceous frustule. The Medusettida (Pls. 118-120) show a peculiar alveolar structure, numerous small compartments being enclosed between two parallel plates. In the Circoporida (Pls. 114-117) and Tuscarorida (Pl. 100) the opaque porcellanous shell has a peculiar cement structure (§ 104), and the lattice-structure is confined for the most part to characteristic rings of pores at the base of the hollow tubes, which arise from the shell. The most peculiar lattice-work, however, appears in the segmented shell of the Aulosphaerida (Pls. 109-111) and Cannosphaerida (Pl. 112). In the former the large meshes of the lattice-work are usually subregular and triangular, in the latter polygonal ; the trabeculae are hollow cylinders, filled with jelly, and containing usually a central axial thread. In each nodal point of the lattice, in which three or more tangential tubes meet, these are separated by stellate or astral septa.
135. Radial Spines of the Skeleton-The skeleton in the great majority of Radiolaria is armed with radial spines, which are of great importance in the development of their general form and of their vital functions. From a morphological point of view the number, arrangement, and disposition of the spines is usually the determining factor as regards the general form of the skeleton. Physiologically they discharge distinct functions, as organs of protection and support ; they act also, like the tentacles of the lower animals, as prehensile organs, since their points, lateral branches, barbed hooks, &c. serve to hold fast nutritive materials. In general main-spines and accessory spines may be distinguished in most Radiolaria ; the former are of pre-eminent importance in determining the figure of the skeleton ; the latter are merely appendicular organs. The main-spines present such characteristic and important differences in the various legions of Radiolaria that they must be considered separately.
136. Radial Spines of the Spumellaria.-The radial spines, which exhibit most manifold variations in the large order Sphaerellaria, present characteristic differences in its four suborders. In the Sphaeroidea their number and disposition serve for the separation into families (p. 59); the Cubosphaerida (Pls. 21-25) always possess six radial main-spines, which stand opposite to each other in pairs and lie in three diameters of the shell, which are at right angles to each other and correspond to the axes of the regular crystallographic system. TheStaurosphaerida (Pl. 15) have four spines, which form a regular cross and stand opposite to each other in pairs, in two axes at right angles. The Stylosphaerida (Pls. 13-17) show only two main-spines, which are opposed to each other in the vertical main axis of the body. Finally, the Astrosphaerida (Pls. 18-20, 26-30) are characterised by a larger and variable number of radial spines (eight, twelve, twenty or more), sometimes regularly, sometimes irregularly arranged. Among the other Sphaerellaria the Prunoidea (Pls. 13-17, 39, 40) are most allied to the Stylosphaerida with two opposite main-spines ; the Discoidea (Pls. 31-47), on the other hand, to the Staurosphaerida with four crossed spines ; there exist, however, Discoidea with two opposite, three marginal, or numerous radial main-spines ; it is characteristic of this suborder that they all usually lie in the horizontal median plane of the lenticular shell, arising from its equatorial margin. The Larcoidea (Pls. 9, 10, 49, 50) show a great variety in the number and arrangement of their radial main spines, which in the different families of this suborder stand in direct causal relation to the various forms of growth of the shell ; usually the primary main-spines lie either in the three different dimensive axes, at right angles to each other, whose differentiation is characteristic of the lentelliptical Larcoid shell (§§ 34, 122) or in definite diagonal axes, which cut the former obliquely. The radial spines of the SPUMELLARIA are never united in the centre of the body, but arise separately from the surface of the primary central lattice-shell (medullary shell), more rarely from one of the secondary (cortical) shells, which enclose it. Their form is originally three-edged (sometimes pyramidal, sometimes prismatic) ; the cause of this is to be found in their origin from the nodal points of the lattice-shell, whose meshes are primitively hexagonal ; hence three trabeculae unite in each nodal point, and are produced into the three edges of the spine. Very commonly, however, the spines are round (conical or cylindrical), more rarely polygonal. The three edges are often delicately toothed, not unfrequently spirally twisted around the axis of the spine (Pl. 21, figs. 1, 12).
137. Radial Spines of the Acantharia.-The radial spines of this legion have a much greater significance than in the other three classes of Radiolaria, since here alone they are the primary determining factors in the skeletal structure, and grow outwards from the middle of the central capsule. This centrogenous origin of the radial spines is as characteristic of the ACANTHARIA as their chemical constitution, which is not siliceous but acanthinic (§ 102). Furthermore, their form is in most cases so peculiar that even an isolated ACANTHARIAN spine can be generally distinguished from one belonging to either of the other three legions. In the great majority of the ACANTHARIA (all Acanthonida and Acanthophracta) twenty radial spines are constantly present, which, disposed according to a definite geometrical law, make up the skeleton (compare § 110 above and p. 717). The twenty spines are generally simply apposed to each other in the centre (either by the surfaces or the edges of their pyramidal base) ; more rarely they are completely united and form a single star-like piece of acanthin (Astrolithium). Very rarely (Acanthochiasma) each two opposite spines are united so that ten diametric bars cross in the middle of the central capsule. Whilst in the great majority of ACANTHARIA these twenty radial spines are present, the small group Actinelida is characterised by the possession of an inconstant, often very large number, sometimes over one hundred. Among these Actinelida are probably to be found the stem-forms of the whole legion. The variously modified spines of the ACANTHARIA may be grouped in three main categories : (1) round (cylindrical or conical) ; (2) four-edged (prismatic or pyramidal) ; (3) two-edged (leaf- or sword-shaped). The latter very commonly bear two opposite transverse processes, the former four crossed ones. By ramification and union of these apophyses arise the lattice-shells of the Acanthophracta (excepting the Sphaerocapsida.).
138. Radial Spines of the Nassellaria. -The radial spines in this legion show as great a variety in their form as in the SPUMELLARIA, and, as in them, are solid, siliceous bars, usually three-edged (prismatic or pyramidal), or round (cylindrical or conical) ; more seldom they are polygonal in section. The great majority of the NASSELLARIA are, however, distinguished by a triradial structure, three primary radial bars diverging from the base of the central capsule (usually from the centre of the porochora) ; there is usually in addition a fourth apical spine, which passes upwards vertically or obliquely on the dorsal aspect of the central capsule. These three or four typical radial spines of the NASSELLARIA may be derived with great probability from the basal tripod of the Plectoidea (Plagoniscus, Plectaniscus, &c., Pl. 91); and since this tripod is very characteristically combined in Cortina and Cortiniscus with the primary sagittal ring of the Stephoidea, the three typical rays may be generally designated "cortinar feet," in contradistinction to the other radial processes of the NASSELLARIAN skeleton. One of the three descending basal feet (" pes caudalis," Pls. 91-95, c) is always unpaired, and lies in the vertical median plane (or sagittal plane), just as does the vertically directed apical spine, which originally forms the dorsal bar of the sagittal ring, and is produced upwards into the "apical horn," marked a on the plates). The other two basal feet are paired, and diverge right and left, forwards and downwards (" pedes pectorales," p.p.). Six-rayed NASSELLARIA, in which three secondary (interradial) feet are intercalated between the three primary (perradial) cortinar feet, are less common than the three-rayed forms. In some groups the number rises still higher, nine, twelve, or even more secondary feet being intercalated between the three primary. Besides, accessory radial spines may be developed on different parts of the shell, which have sometimes a definite relationship to the typical radial spines, sometimes not. Their form and ramification are very various (Pls. 51-98).
139. Radial Spines of the Phaeodaria.-The radial spines
of the PHAEODARIA are very clearly distinguished from those of other Radiolaria
by the fact that they are usually blow tubes, rarely solid bars. As a rule,
the tubes are cylindrical, often slightly fusiform or conical, their siliceous
wall is very thin, and their lumen filled with jelly ; a fine thread
of silica usually runs in the axis, and in several families is connected
by fine transverse threads with the wall of the tube (Pl. 110,
figs. 4, 6 ; Ph. 115, figs. 6,
7). The peculiar family Medusettida is characterised by a very remarkable
segmentation of the hollow spines (Pls. 118-120).
Each tube is divided by a series of septa into chambers, which communicate
by a central or excentric opening in each septum, an arrangement resembling
the siphuncle of the chambered Cephabopod shells. The number and arrangement
of the radial tubes in most PHAEODARIA is indefinite and very variable
; only in a few families is the number constant in each species and genus,
and the disposition regular. The Medusettida (Pls. 118-120)
resemble the NASSELLARIA, inasmuch as equal radial feet diverge from the
base of their shell, sometimes three in number (Cortinetta, Pl.
117, fig. 9), sometimes four (Medusetta
Pl. 120 figs. 1-4),
sometimes six
(Gazelletta);Gorgonetta
is specially distinguished
by the possession of six ascending and six descending spines regularly
alternating (Pl. 119). The Tuscarorida
(Pl. 100) usually have three or
four equidistant feet. The Circoporida (Pls. 115-117),
on the other hand, rather approach the Sphaeroidea, their spherical or
regular polyhedral shell having a definite number of tubular radial spines,
which arise at regular intervals from their angles ; Circoporus
has
six,
Circospathis
nine,
Circogonia
twelve, and Circorrhegma
twenty
radial tubes. Very rarely the tubes of the PHAEODARIA are angular, usually
they are round, more or less cylindrical, though they are often bifurcated
or even ramified, and exhibit a great wealth of the most delicate appendages
; siliceous hairs, bristles, spines, barbed or anchor-like hooks, spathilke,
brushes, circlets, &c. (compare Pls. 99-128).
140. Main-Spines and Accessory Spines.-As accessory spines (Paracanthae) we oppose to the main-spines (Protacanthae), just described, all those processes which have no determining influence upon the formation of the skeleton as a whole, but are to be regarded as secondary constituents of the skeleton, or appendicular organs of inferior significance. They are developed in the utmost variety, sometimes as hairs or bristles, sometimes as thorns or clubs, either straight or curved (often zigzag), smooth or barbed ; sometimes standing vertically upon the shell, or directed towards the centre, sometimes obliquely, or rising at a definite angle. In those SPUMELLARIA whose lattice-shell consists of several concentric spheres, the accessory spines generally arise from the outermost, the main-spines, on the contrary, from the innermost. In the NASSELLARIA, multifarious forms of accessory spines are especially developed in the order Plectellaria. In the PHAEODARIA they are often furnished with delicate appendages, e.g., anchor-hooks, spathillae, coronets, &c. Among the ACANTHARIA the accessory spines which arise from the surface of the shell in the Acanthophracta are very characteristic. They are not radially disposed (like the similar superficial spines of the SPUMELLARIA), but parallel to the radial main-spines from whose transverse processes they arise. Since in all these Acanthophracta the twenty radial main-spines are opposite to each other in pairs, all the accessory spines (often several hundred) are parallel to ten different regularly disposed axes of the lattice-shell (Pls. 134-138).
The skeletons of the Radiolaria, in addition to the general relations which have been discussed above, present numerous and important special differences in the various larger and smaller groups. These are indicated in detail in the descriptions of the legions, orders, and families in the systematic portion of this Report.
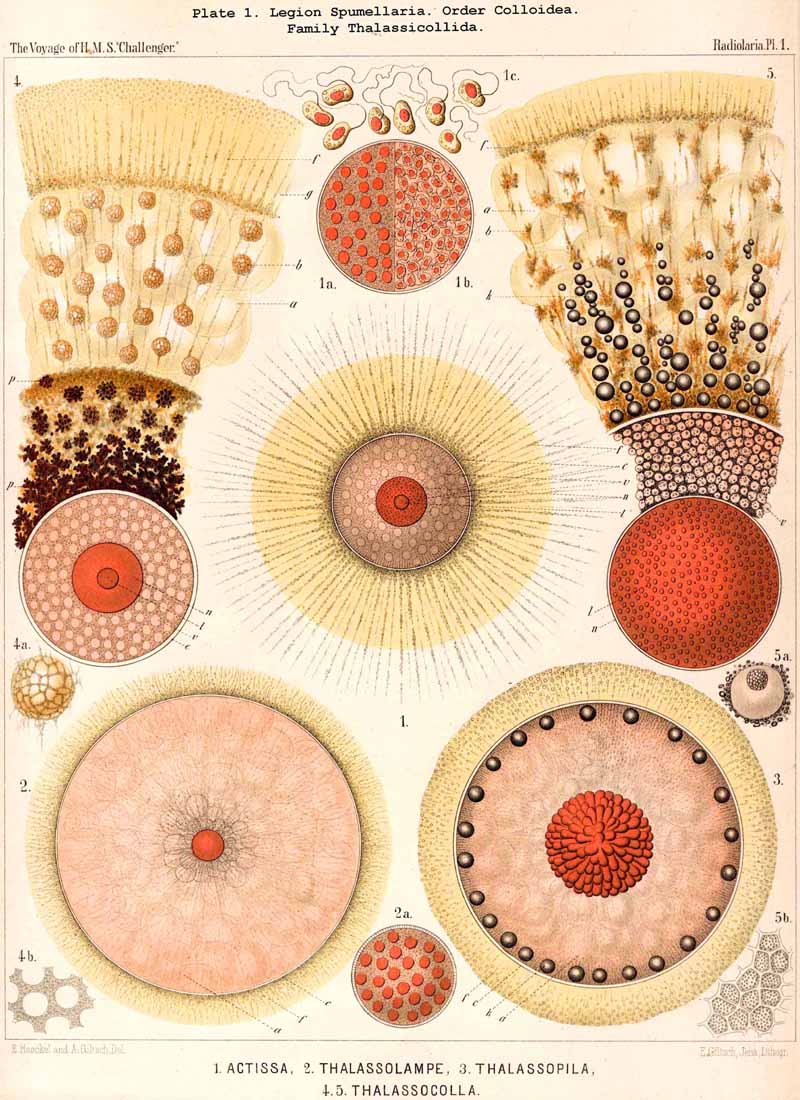{kind=link}
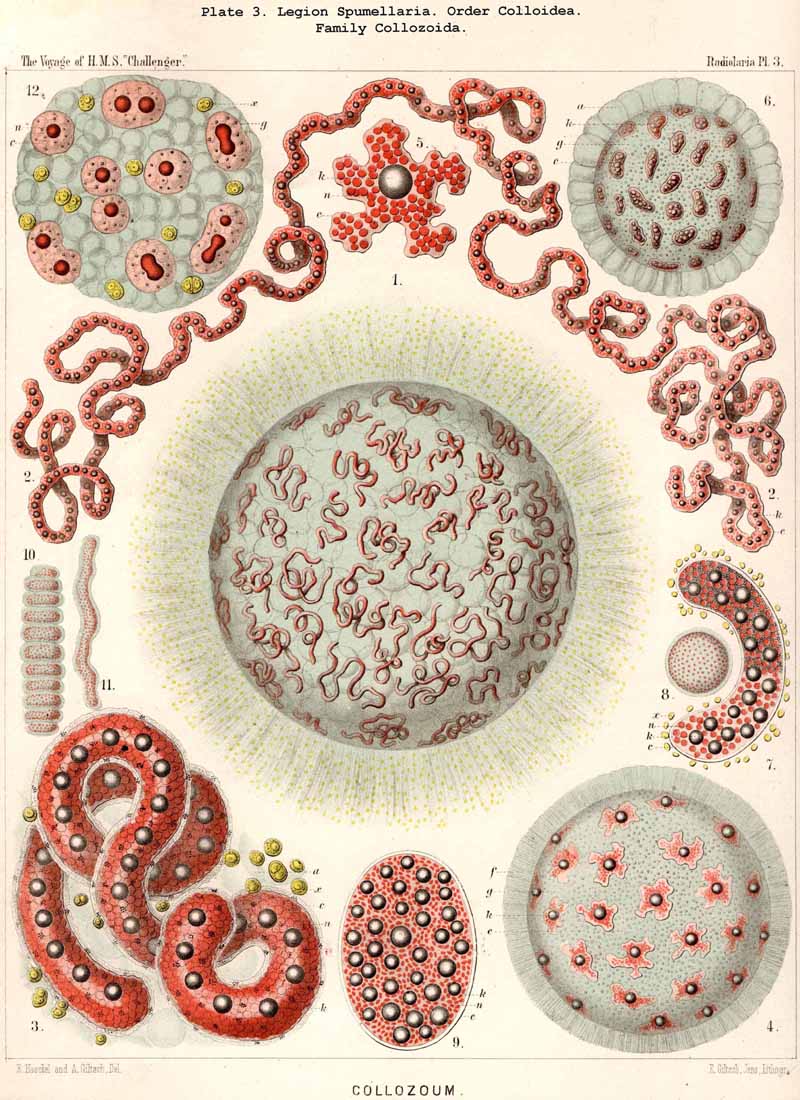{kind=link}
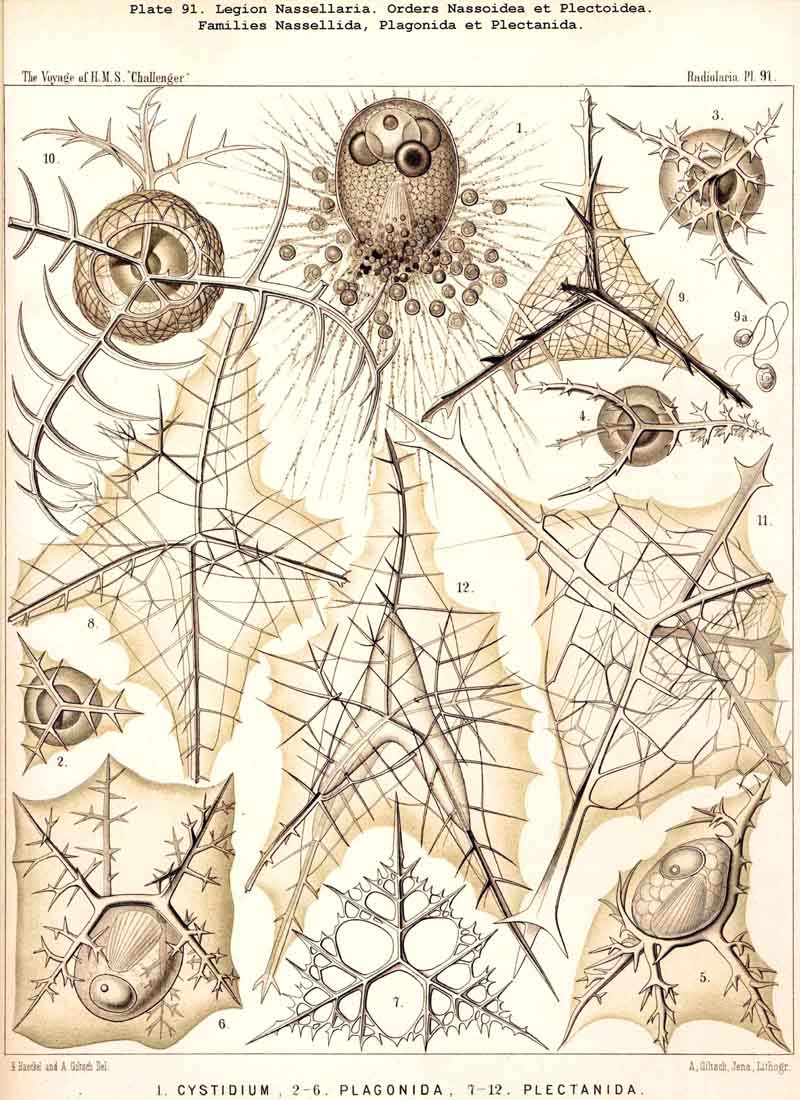{kind=link}
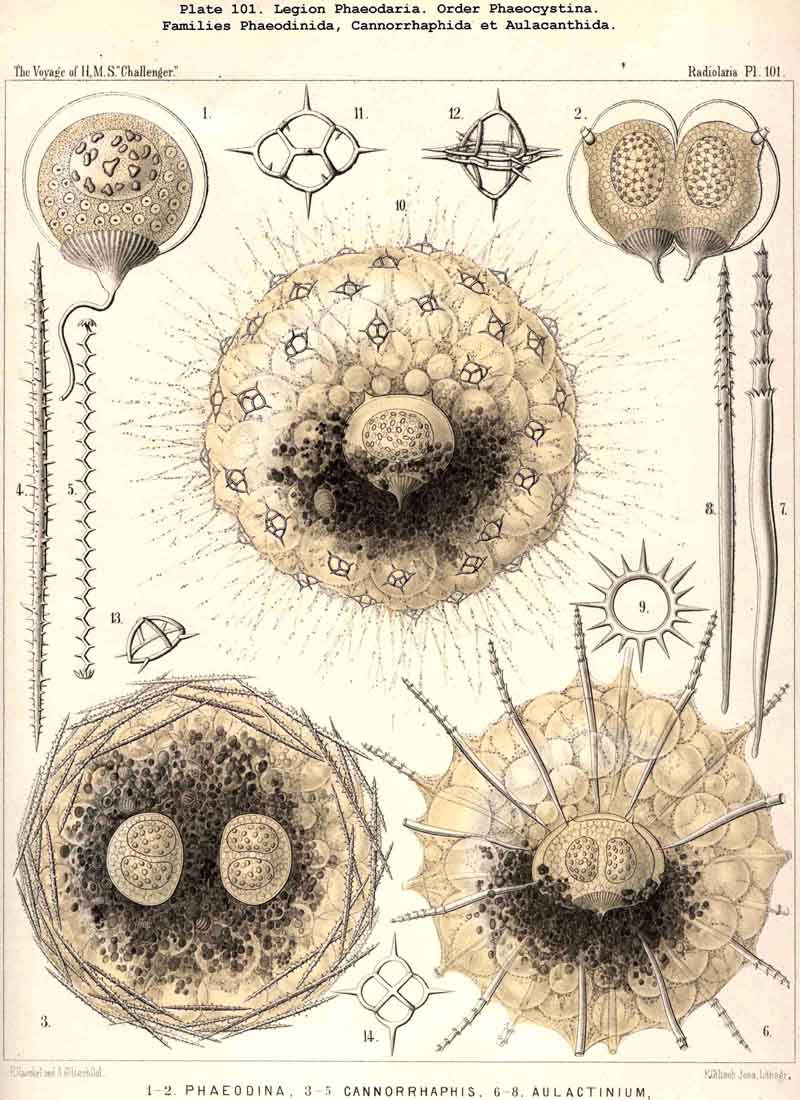{kind=link}
{kind=link}
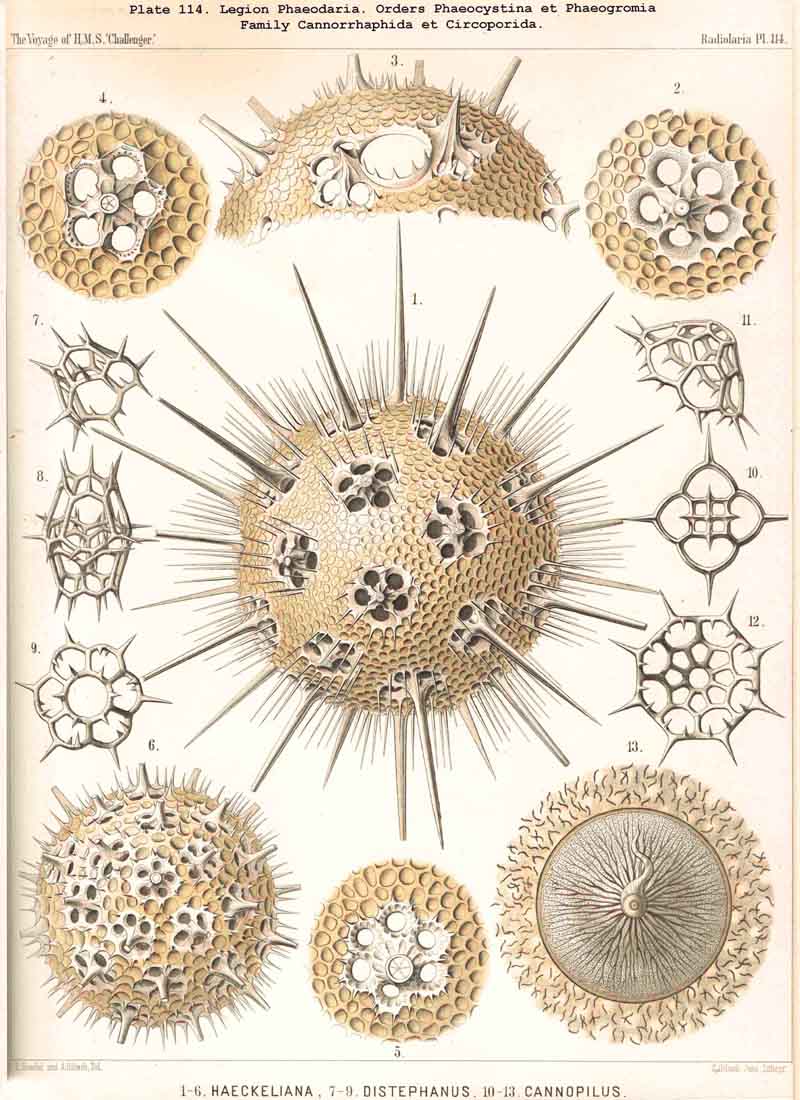{kind=link}
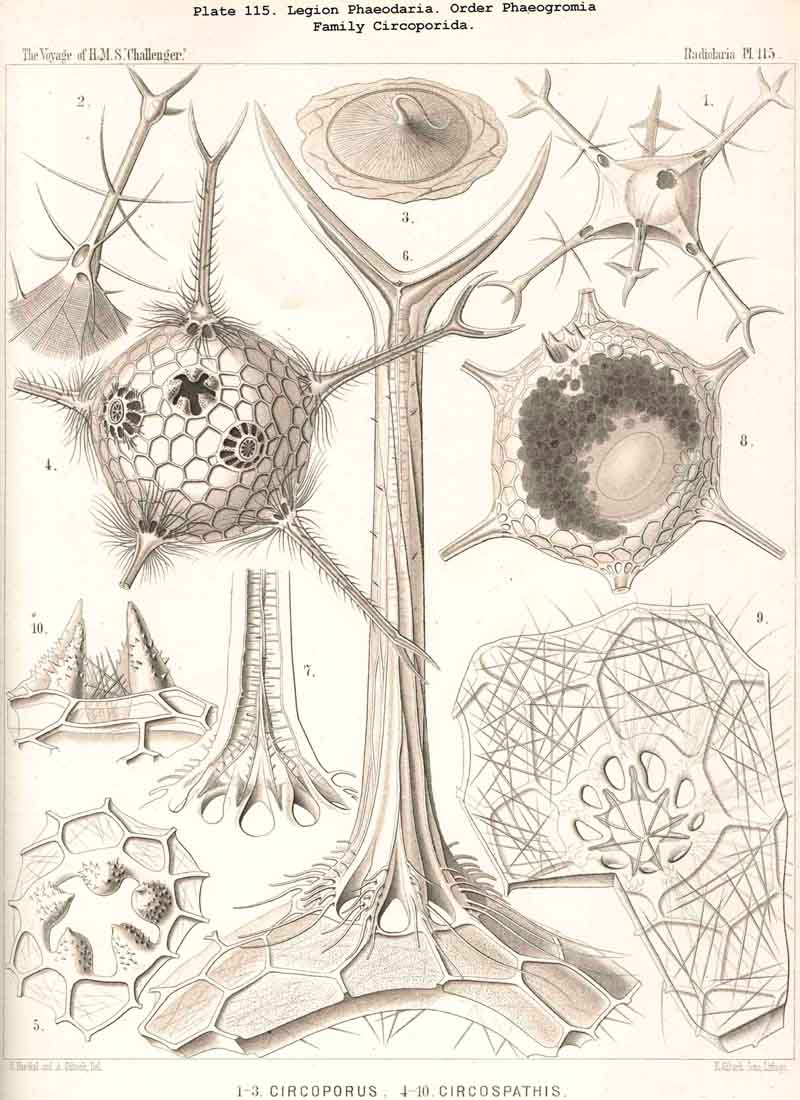{kind=link}
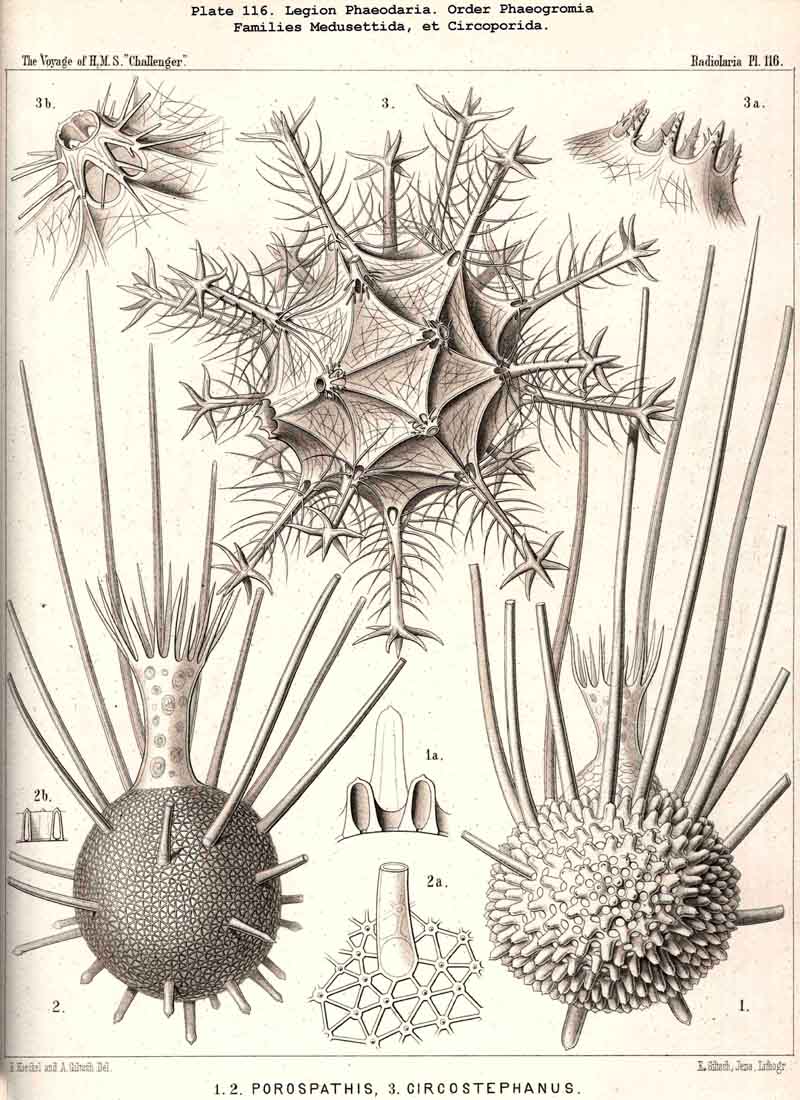{kind=link}
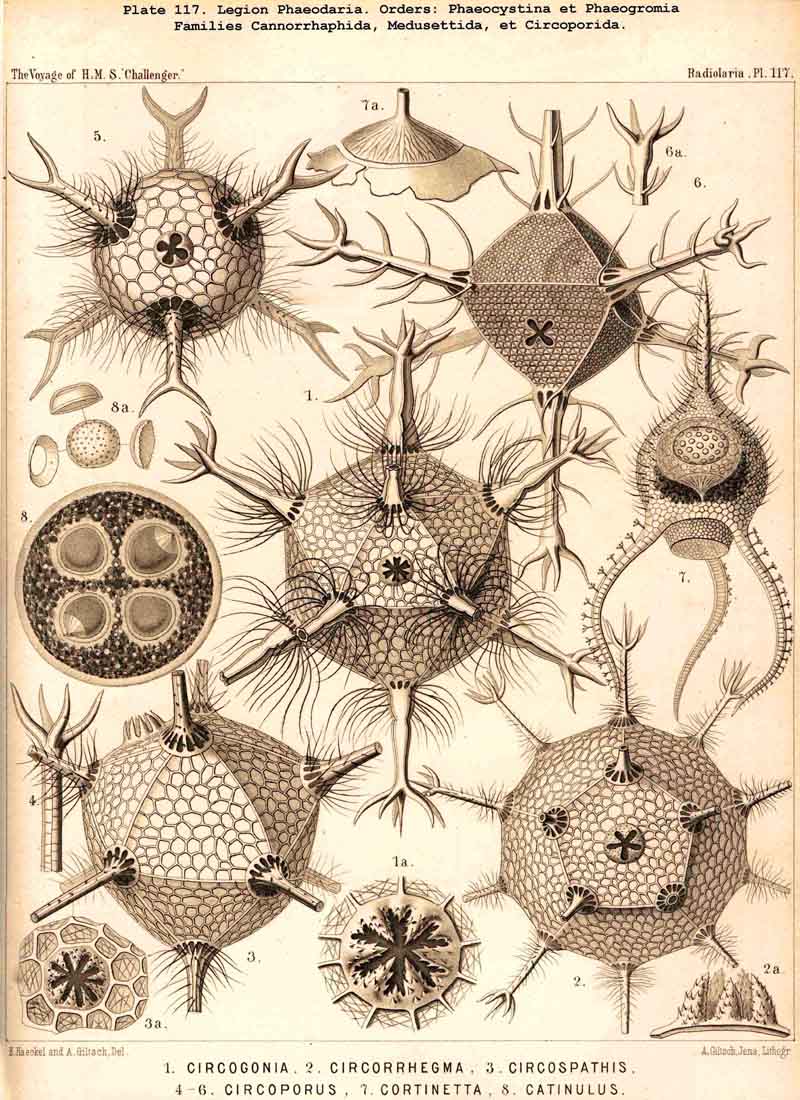{kind=link}
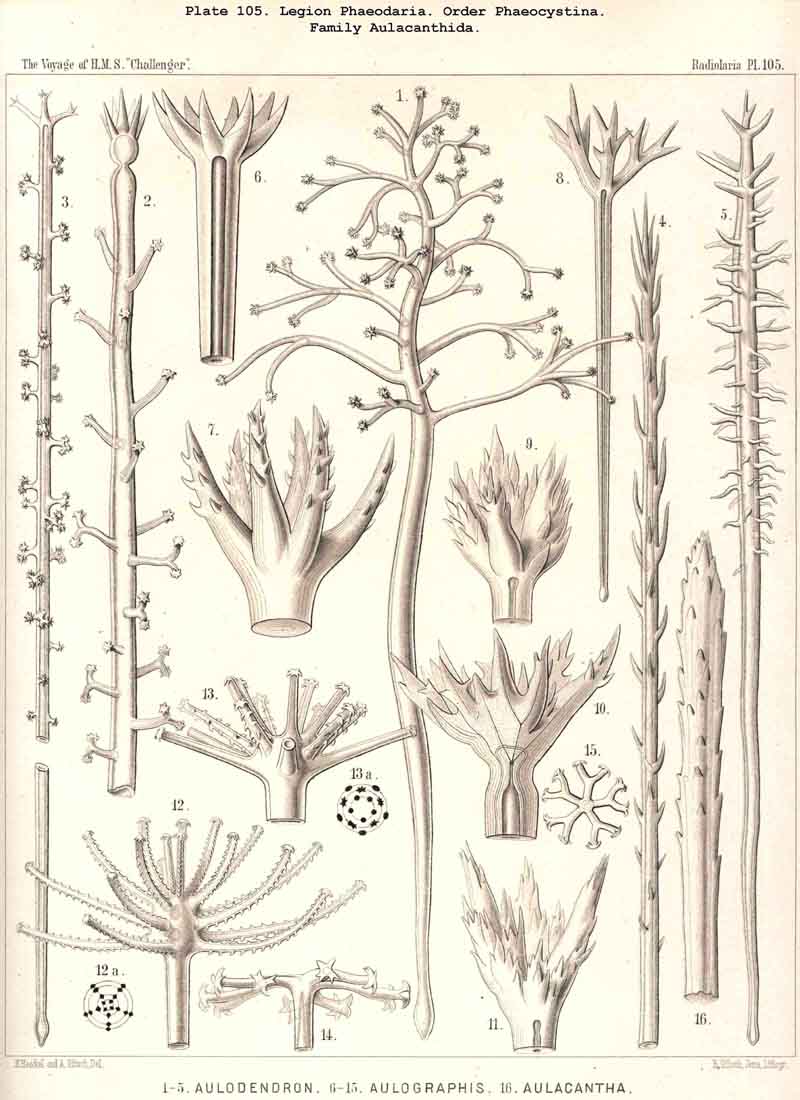{kind=link}
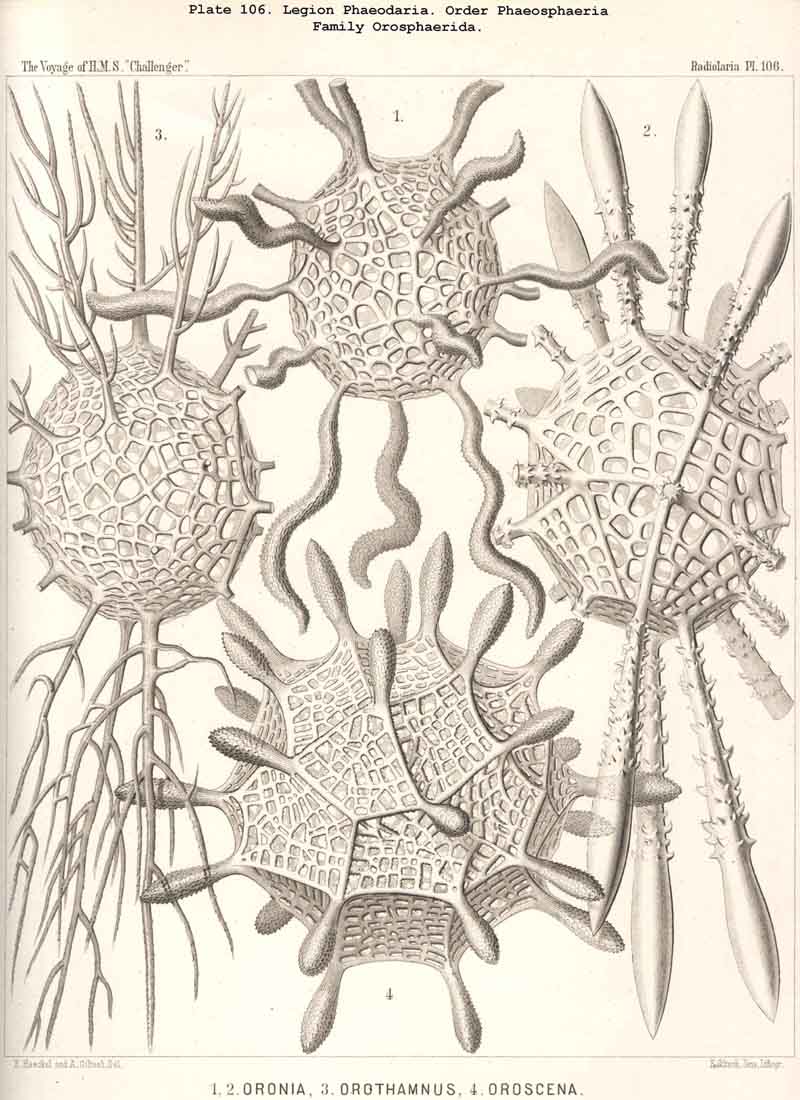{kind=link}
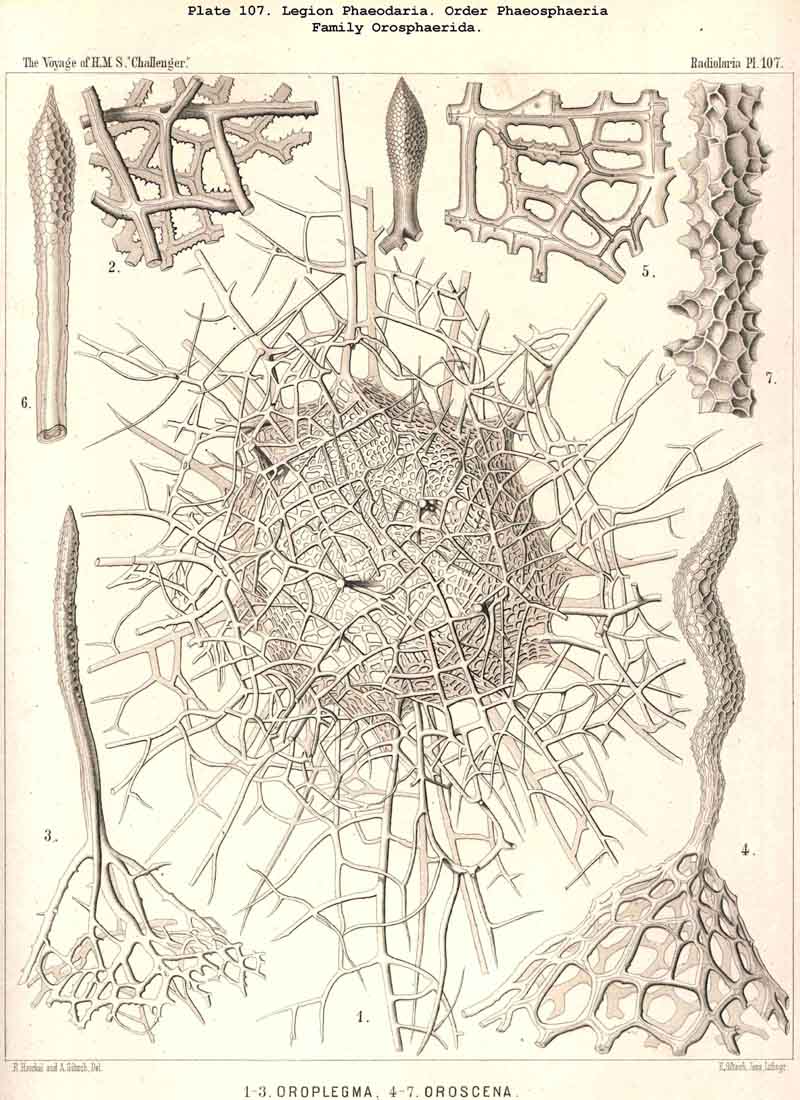{kind=link}
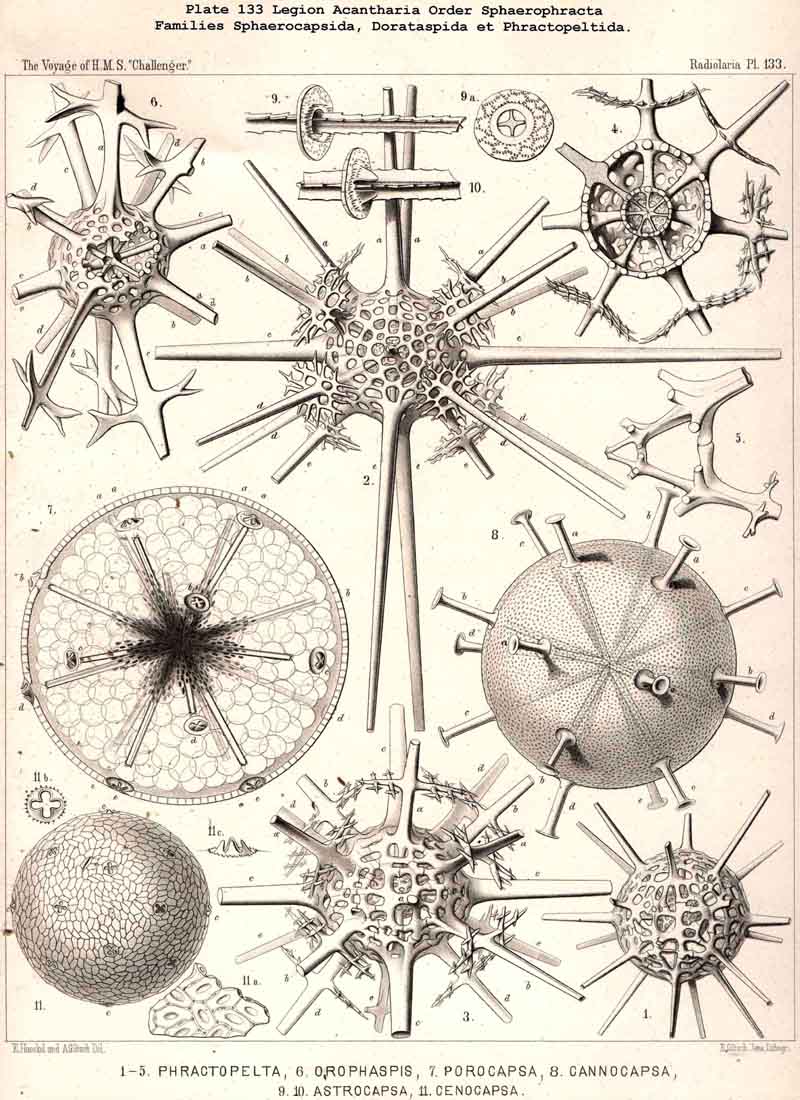{kind=link}
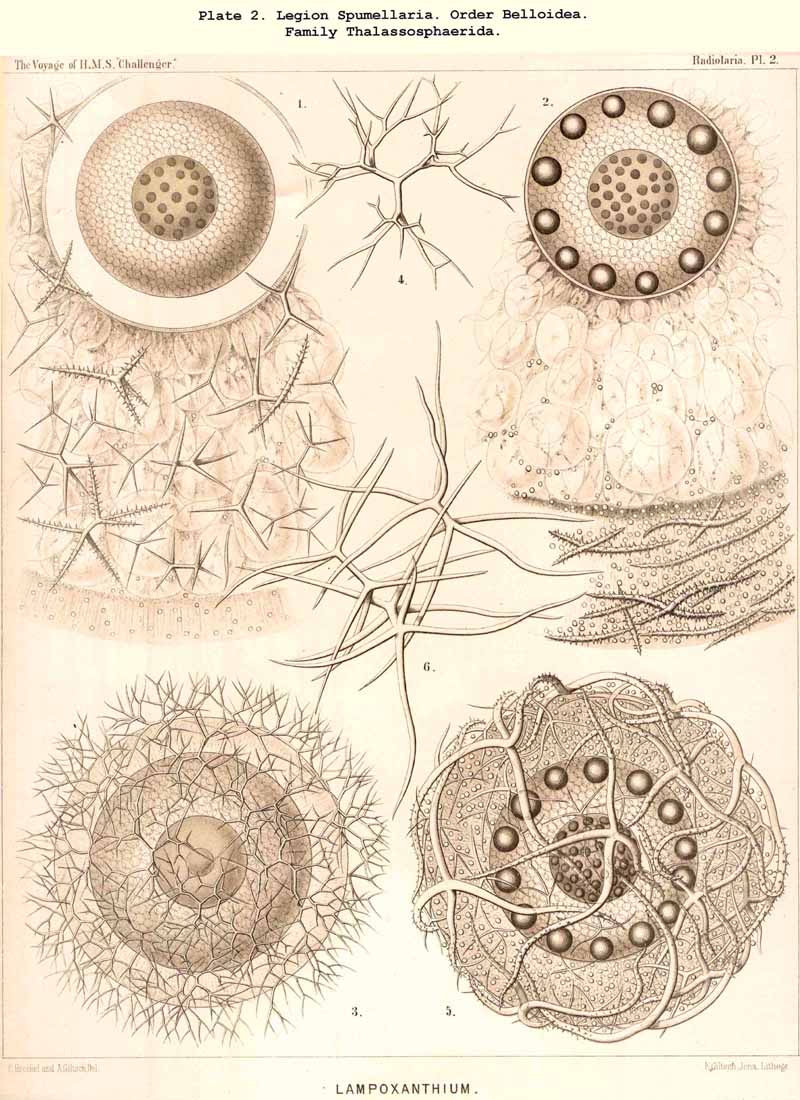{kind=link}
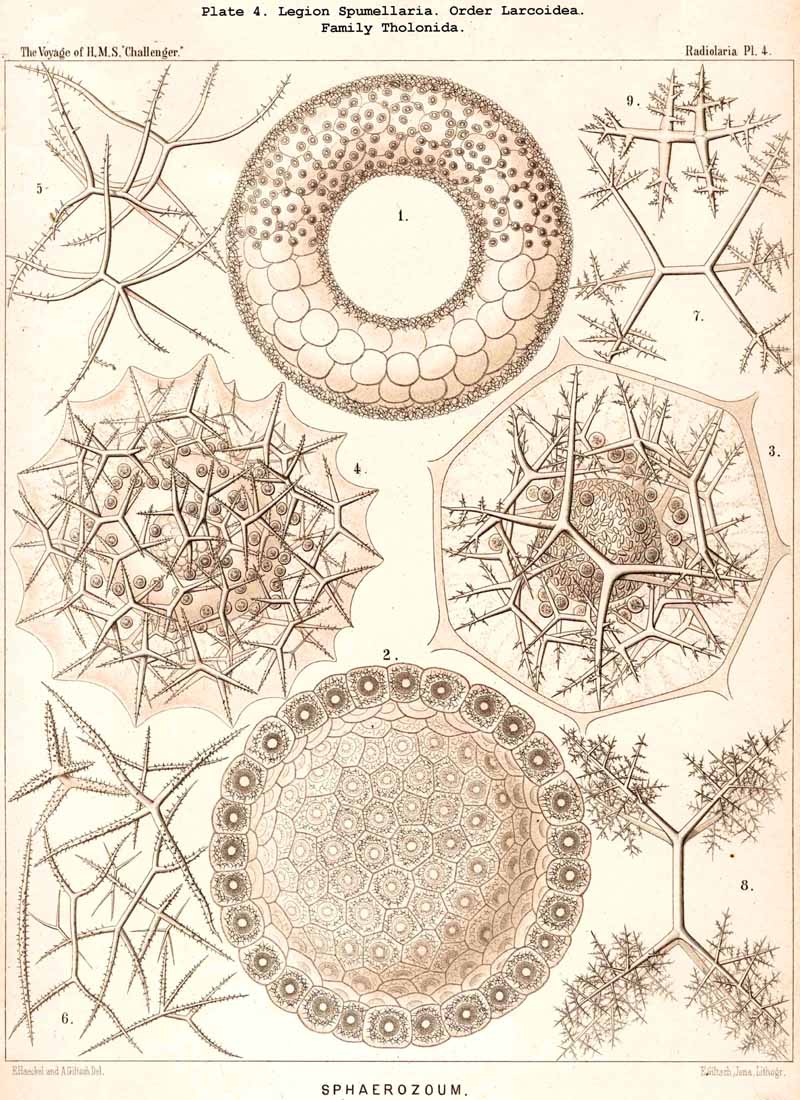{kind=link}
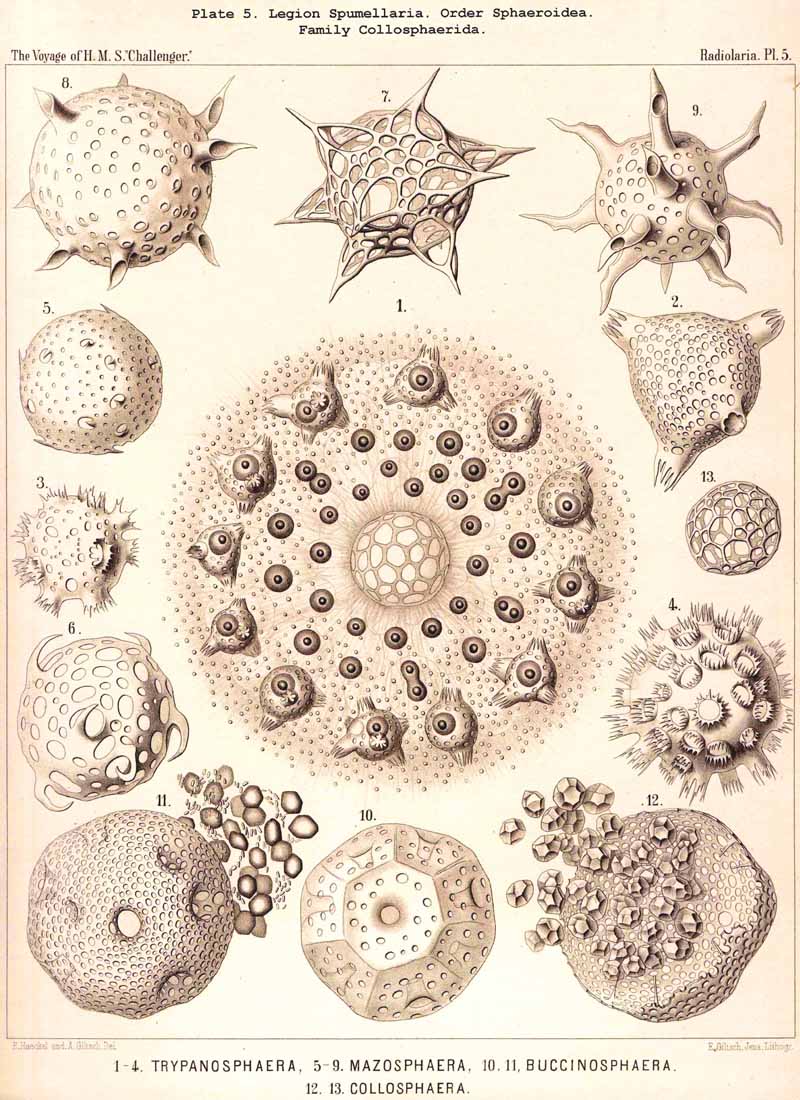{kind=link}
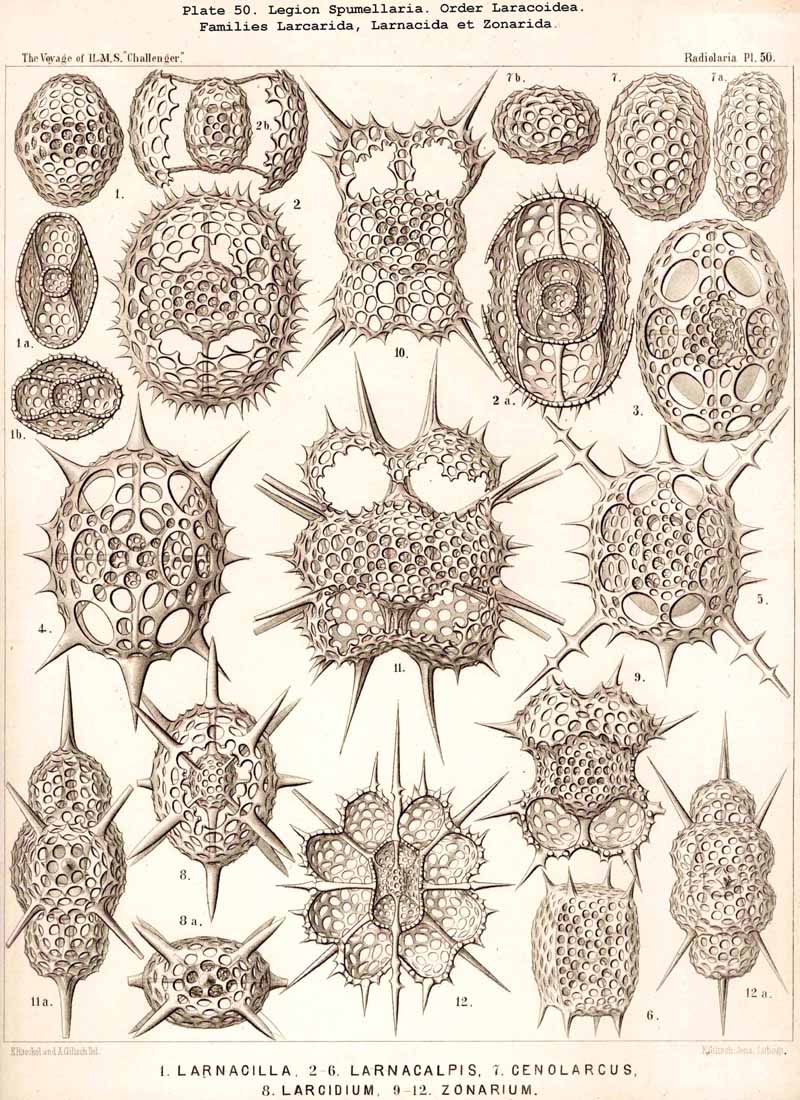{kind=link}
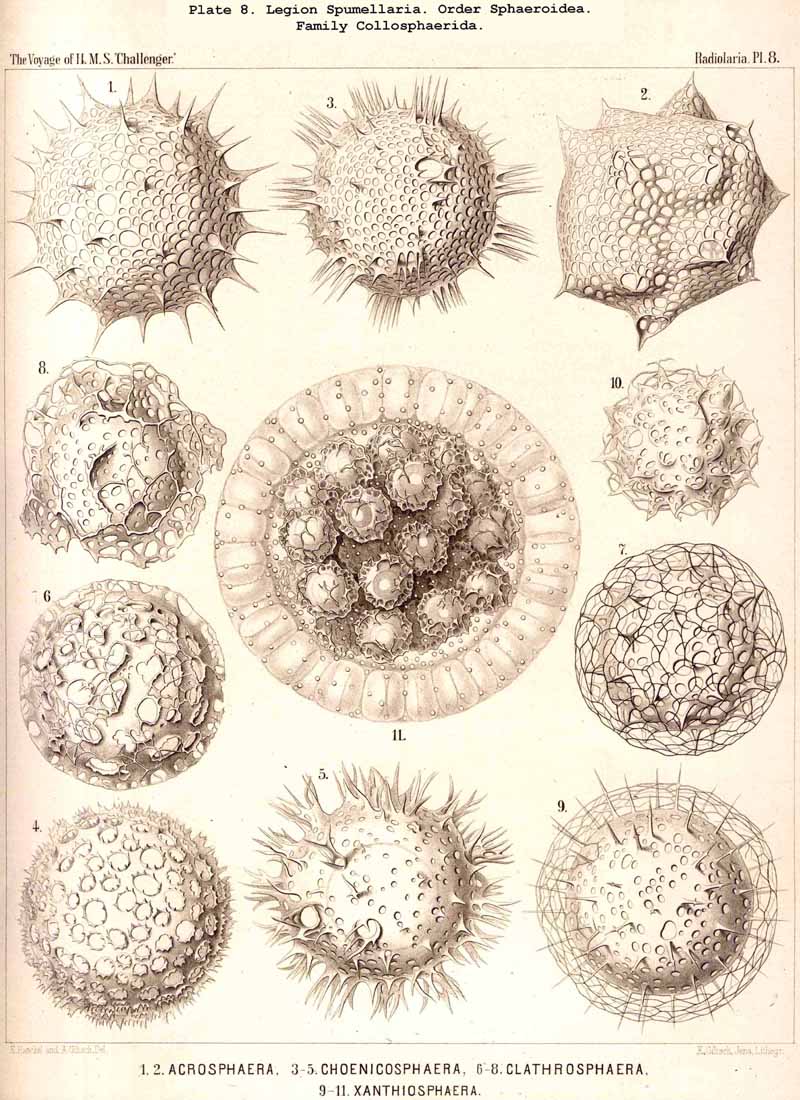{kind=link}
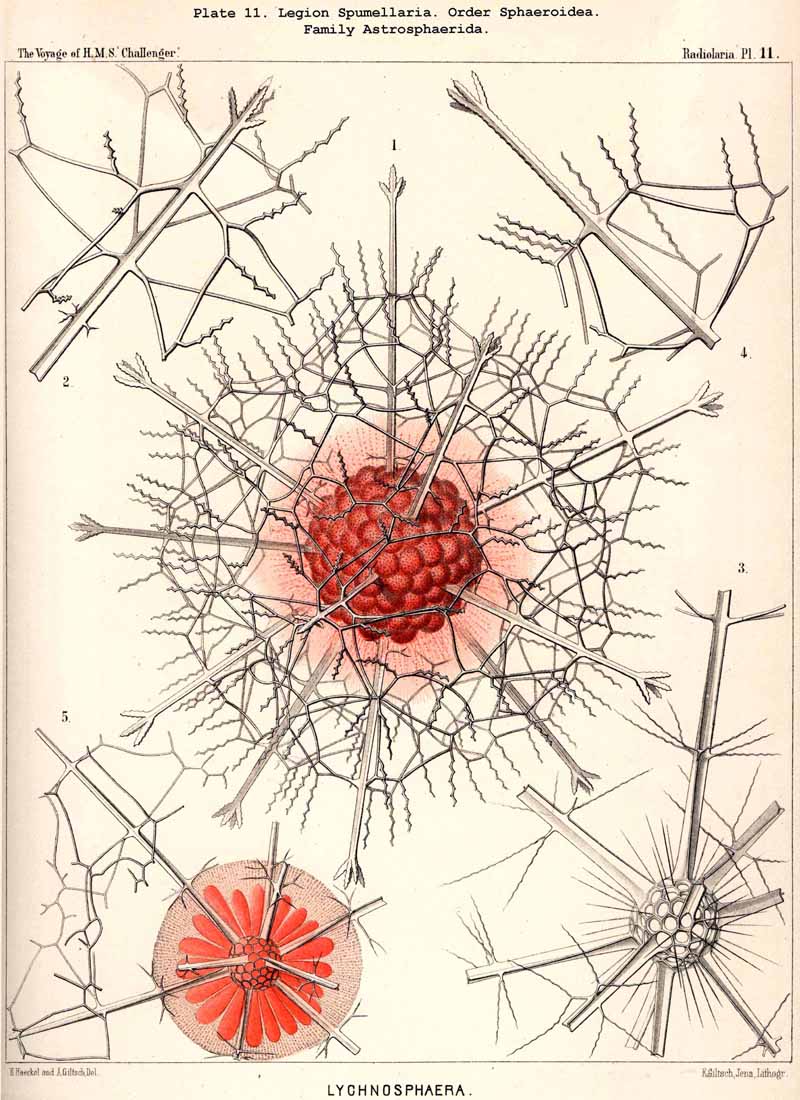{kind=link}
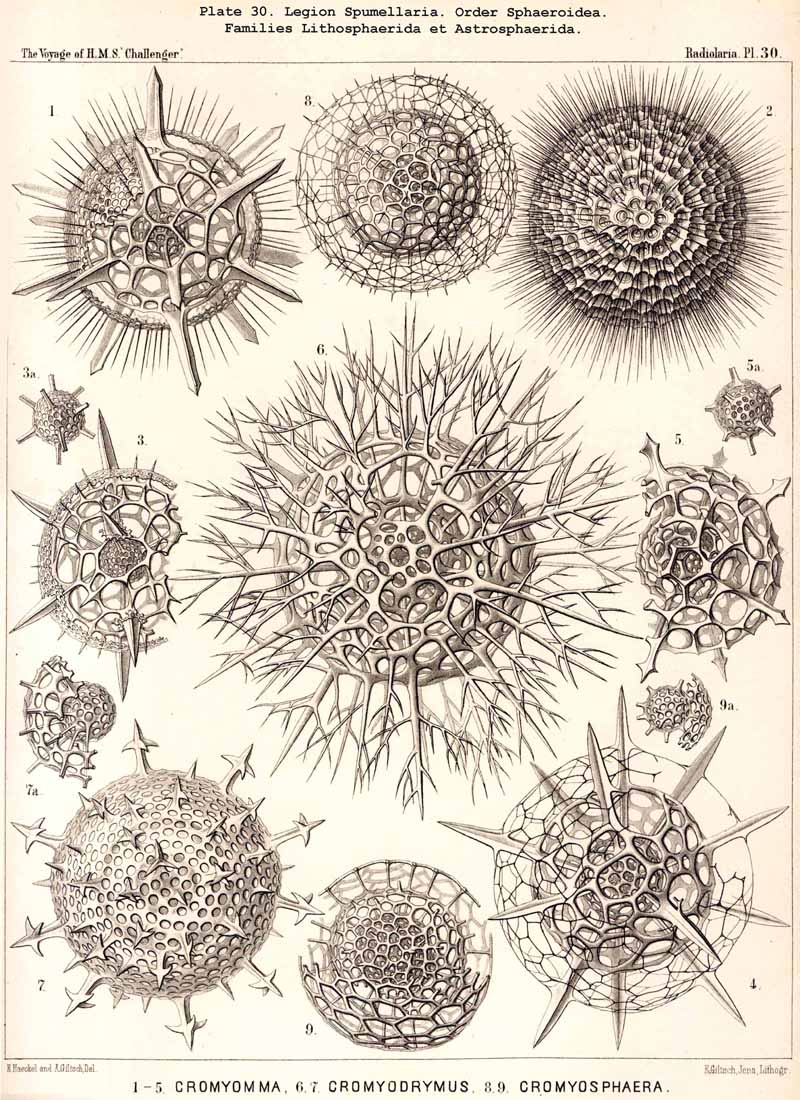{kind=link}
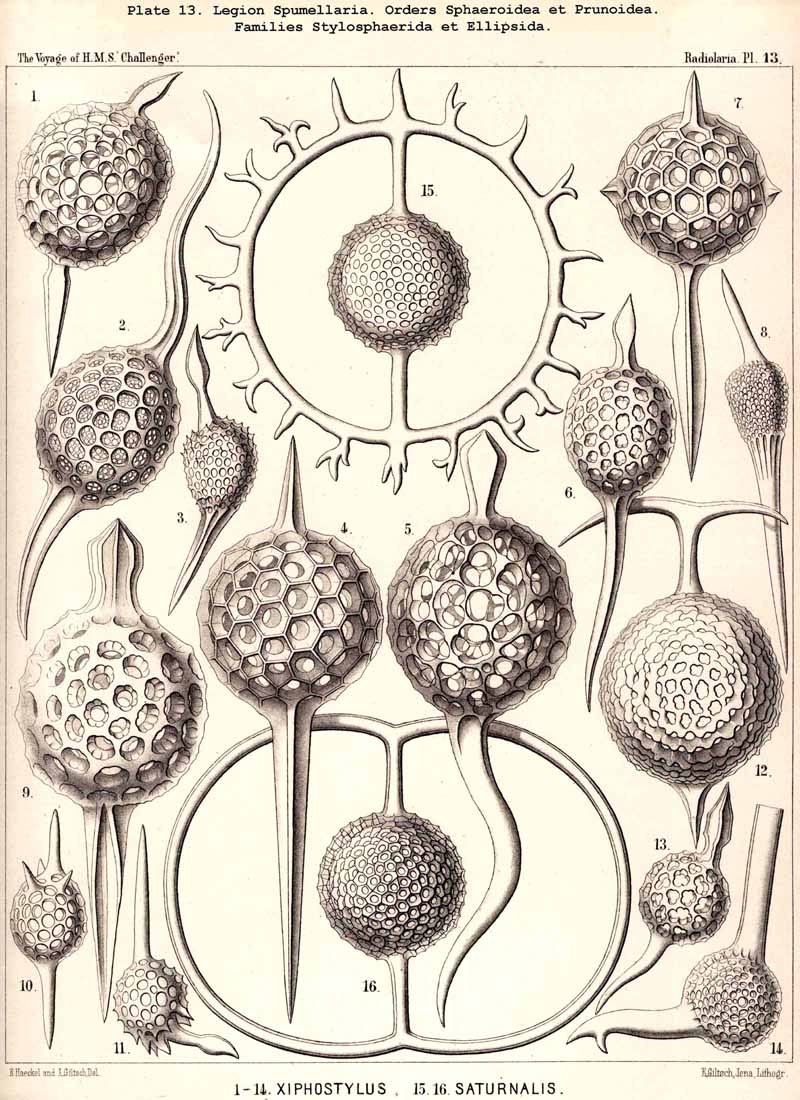{kind=link}
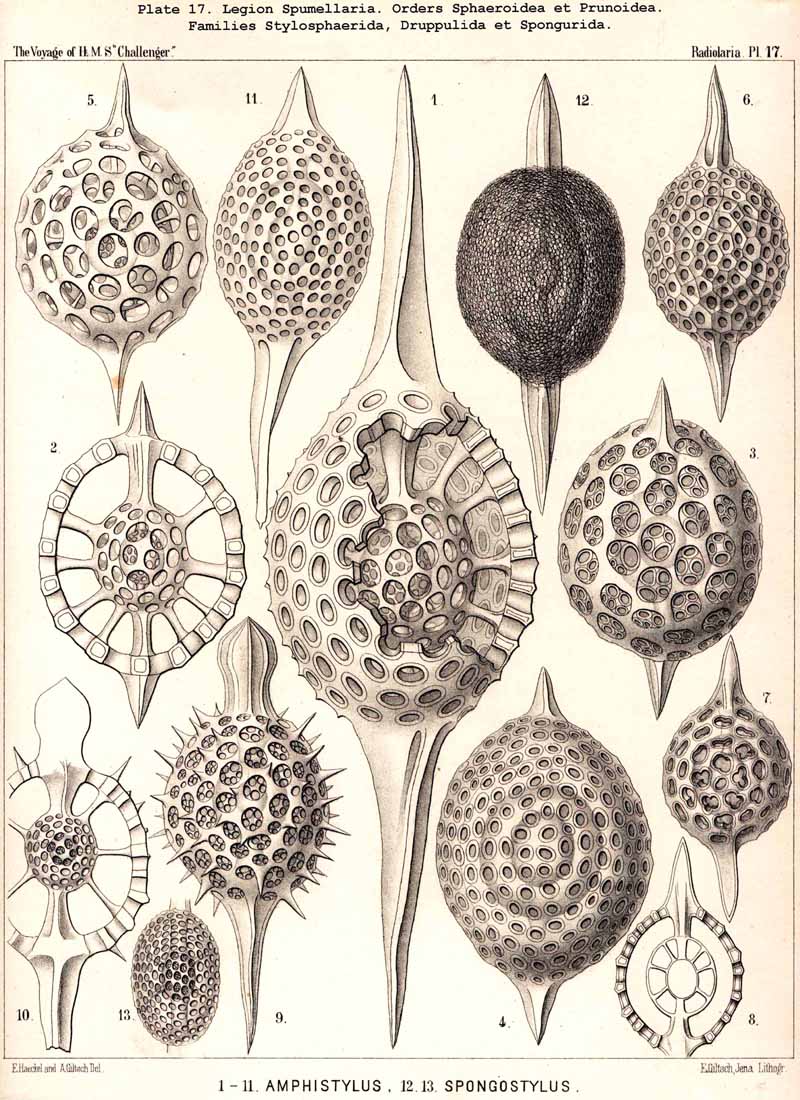{kind=link}
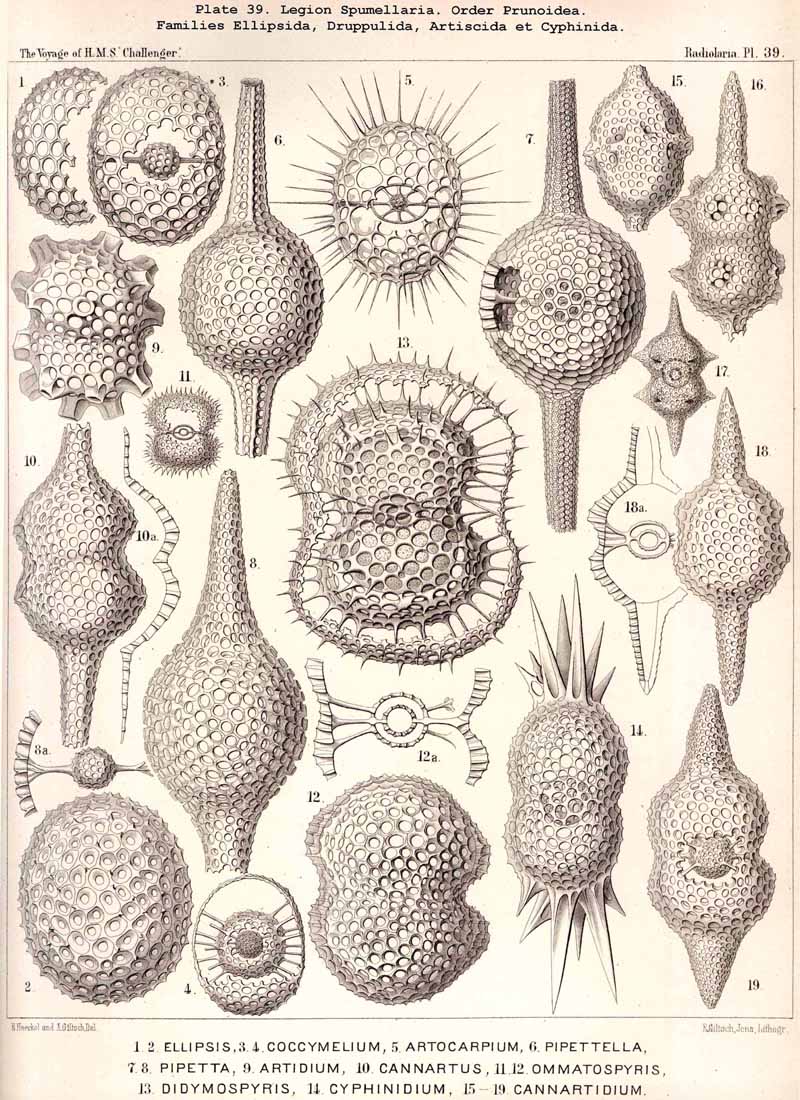{kind=link}
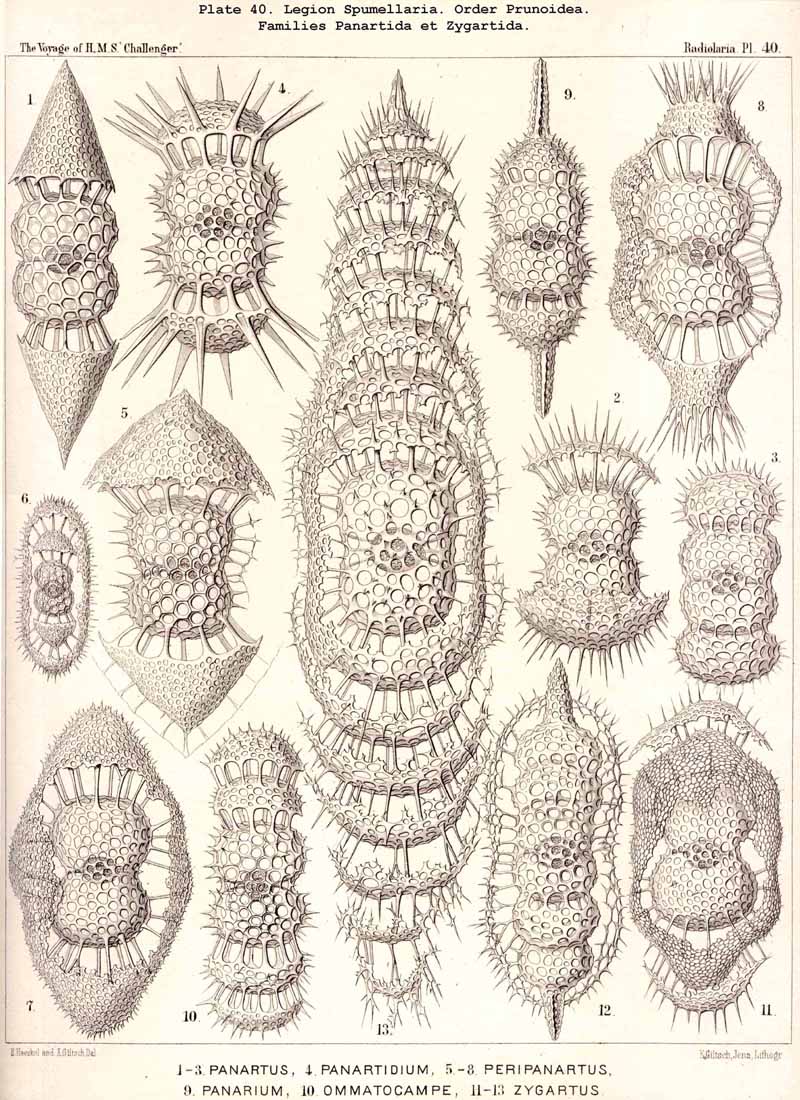{kind=link}
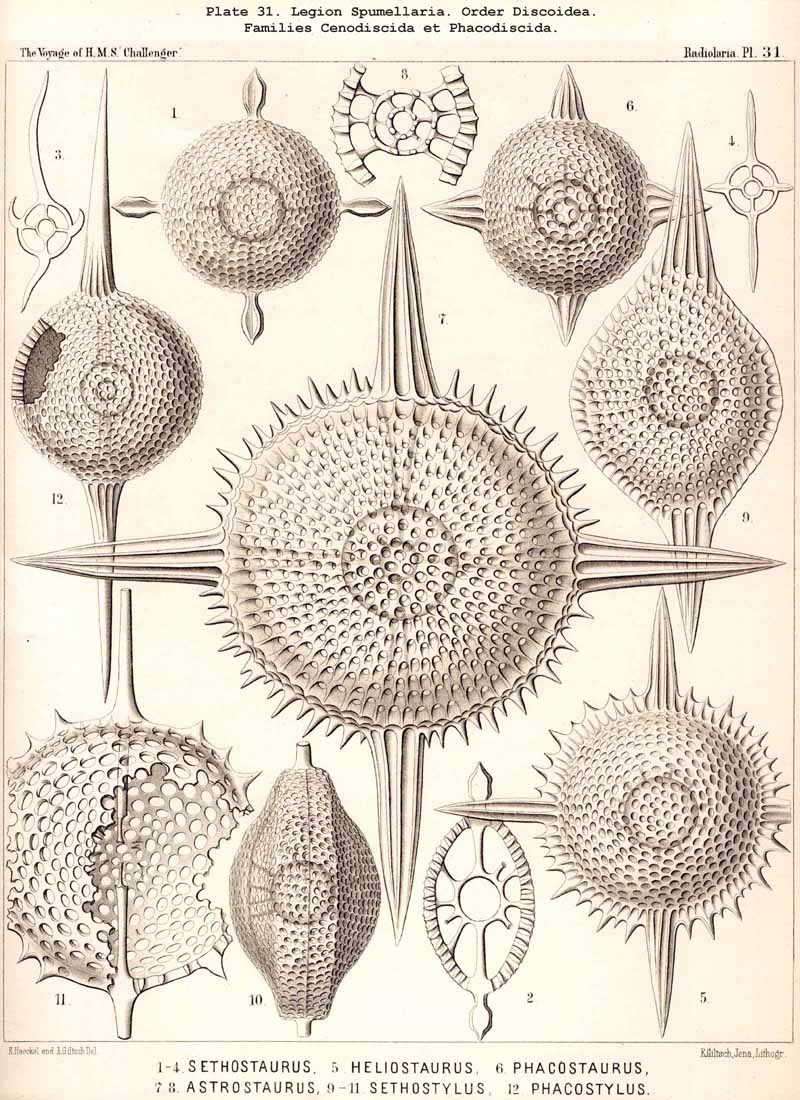{kind=link}
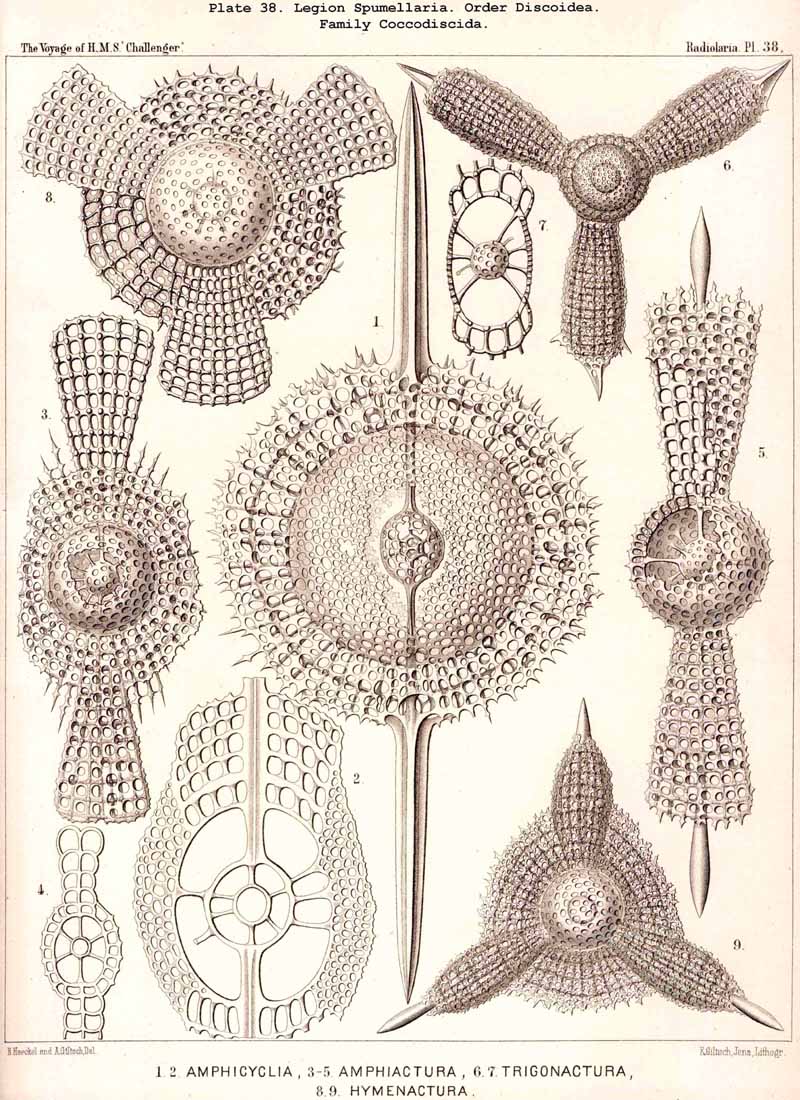{kind=link}
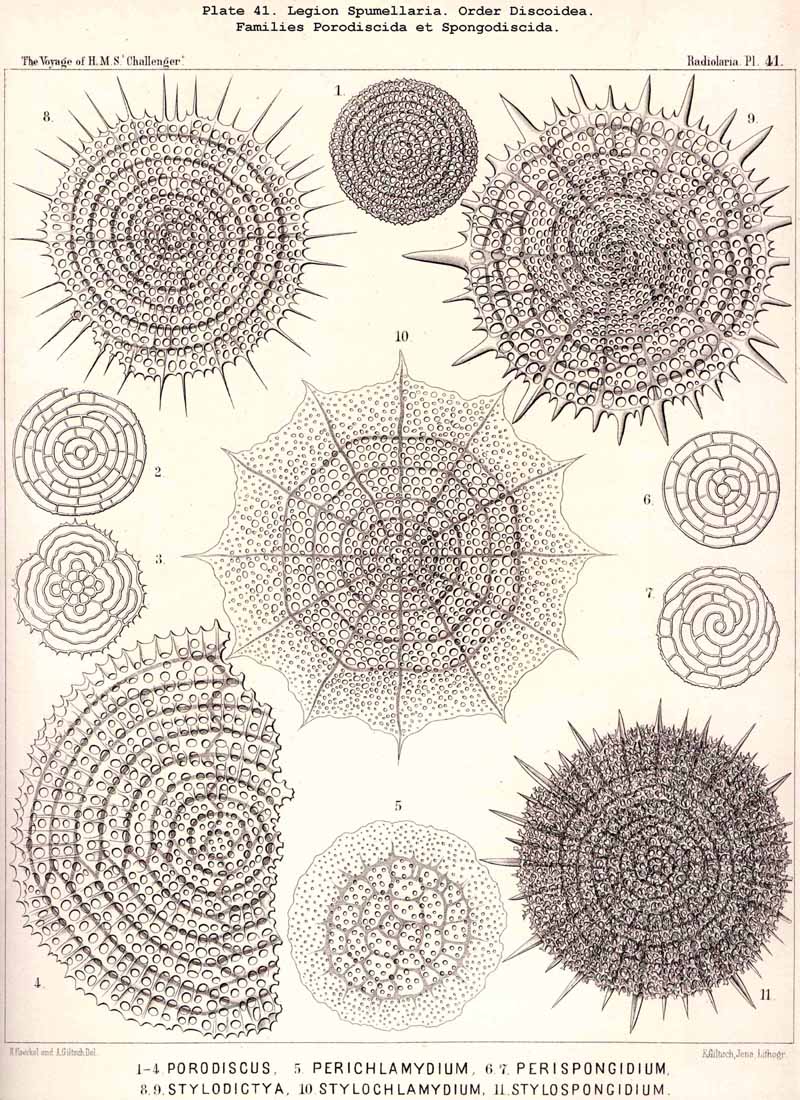{kind=link}
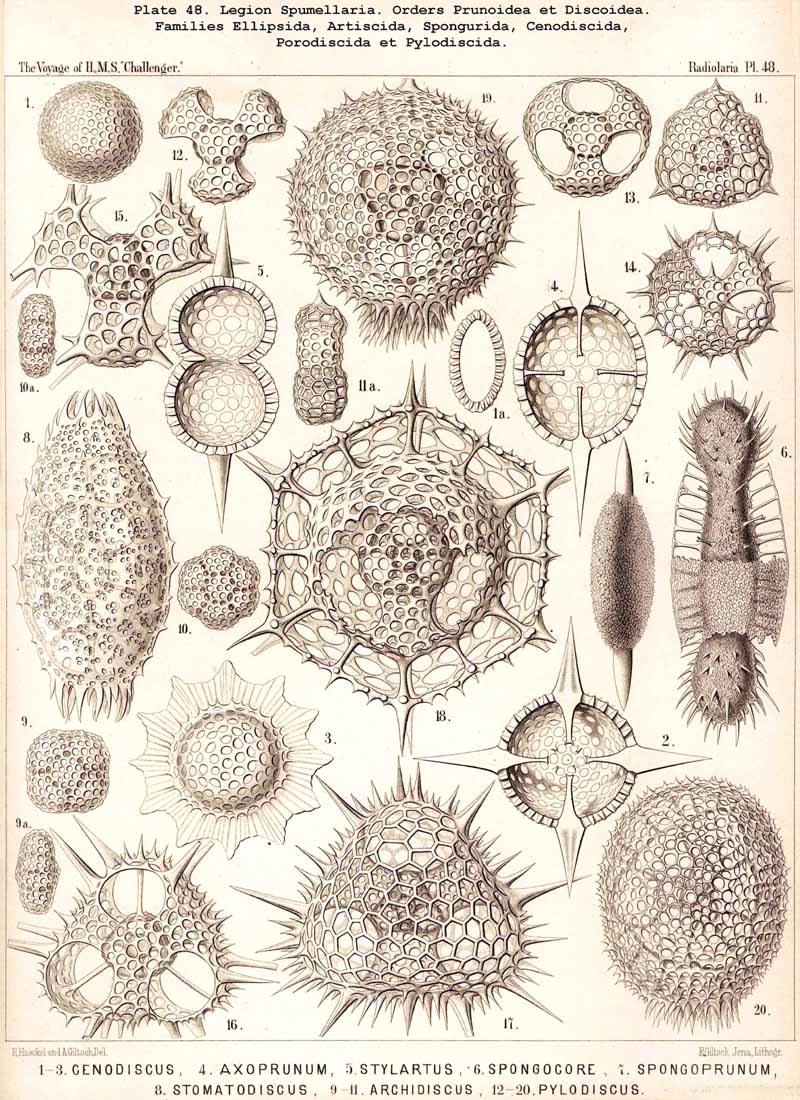{kind=link}
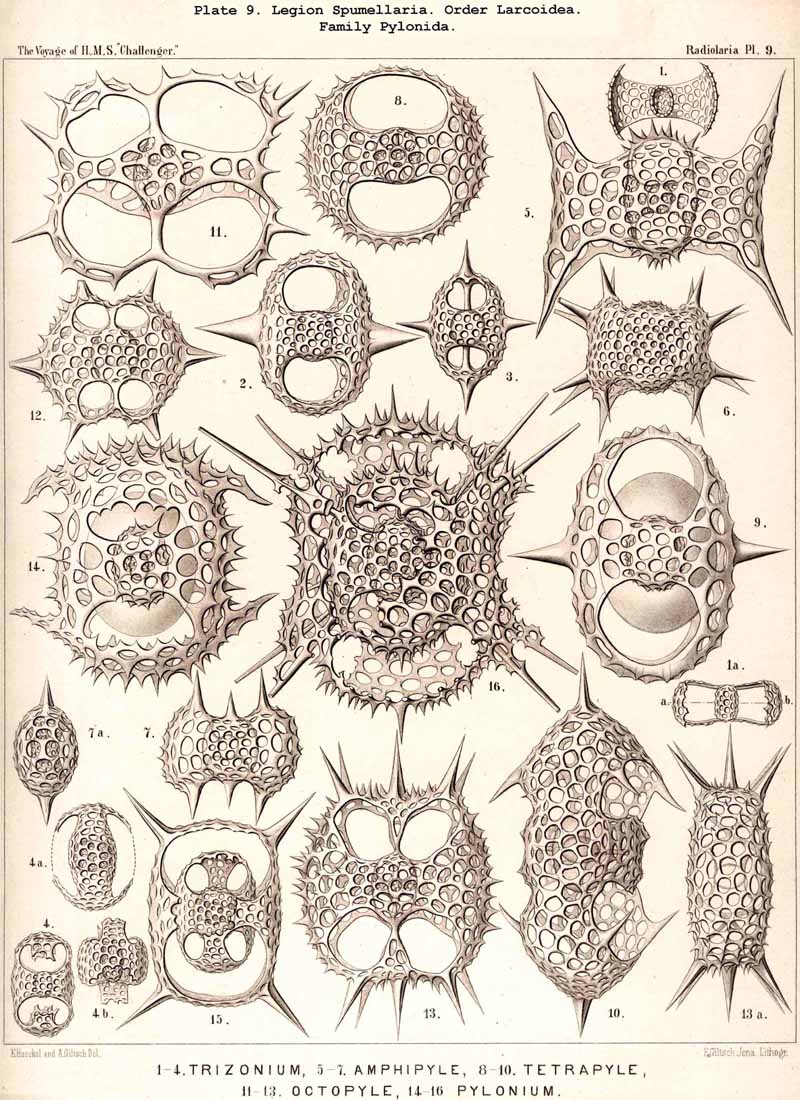{kind=link}
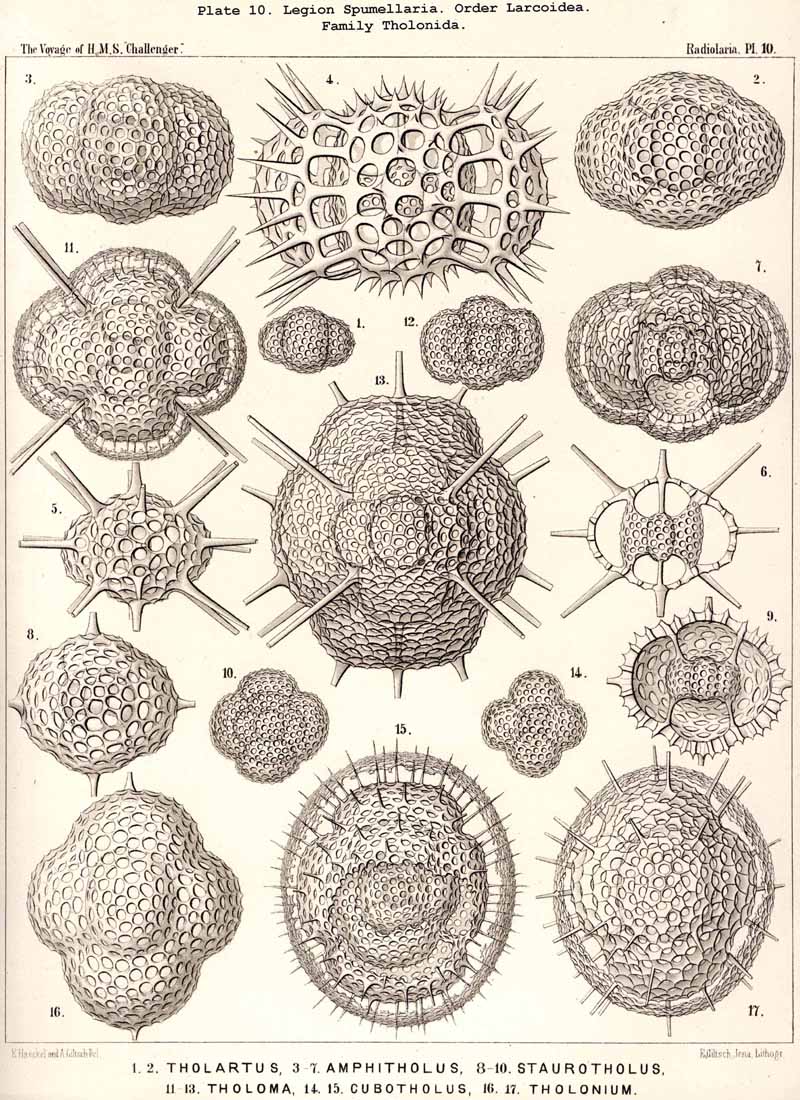{kind=link}
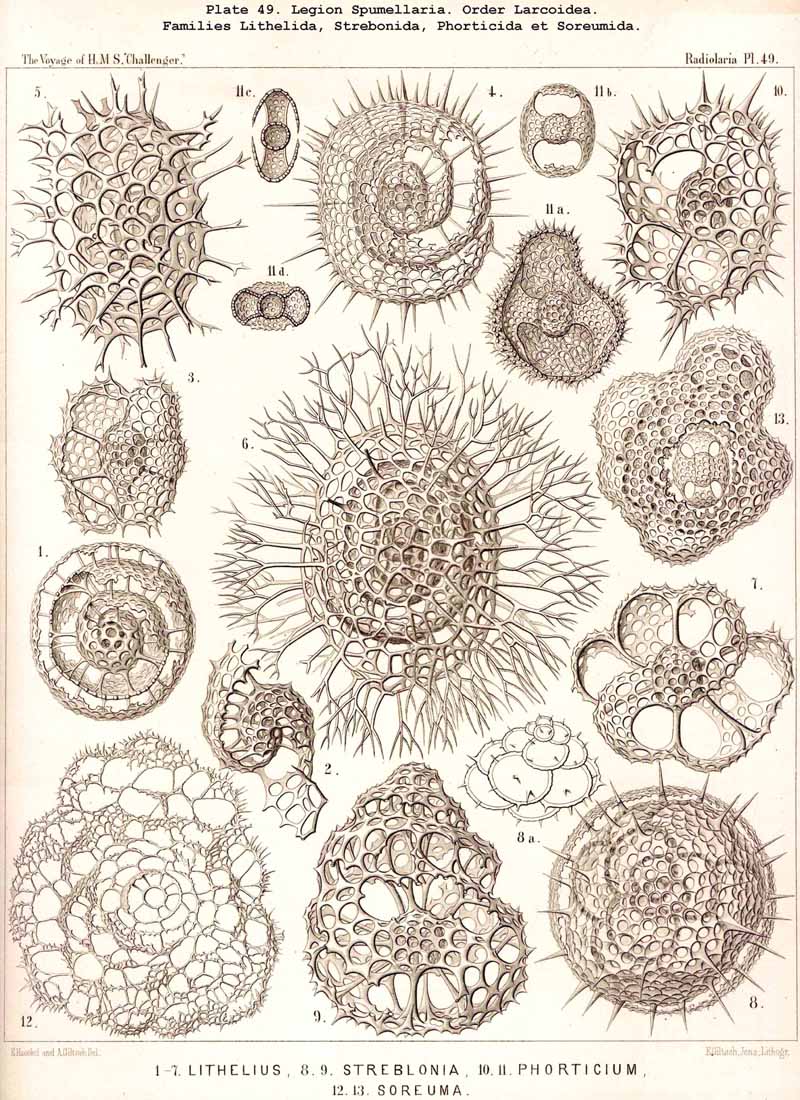{kind=link}
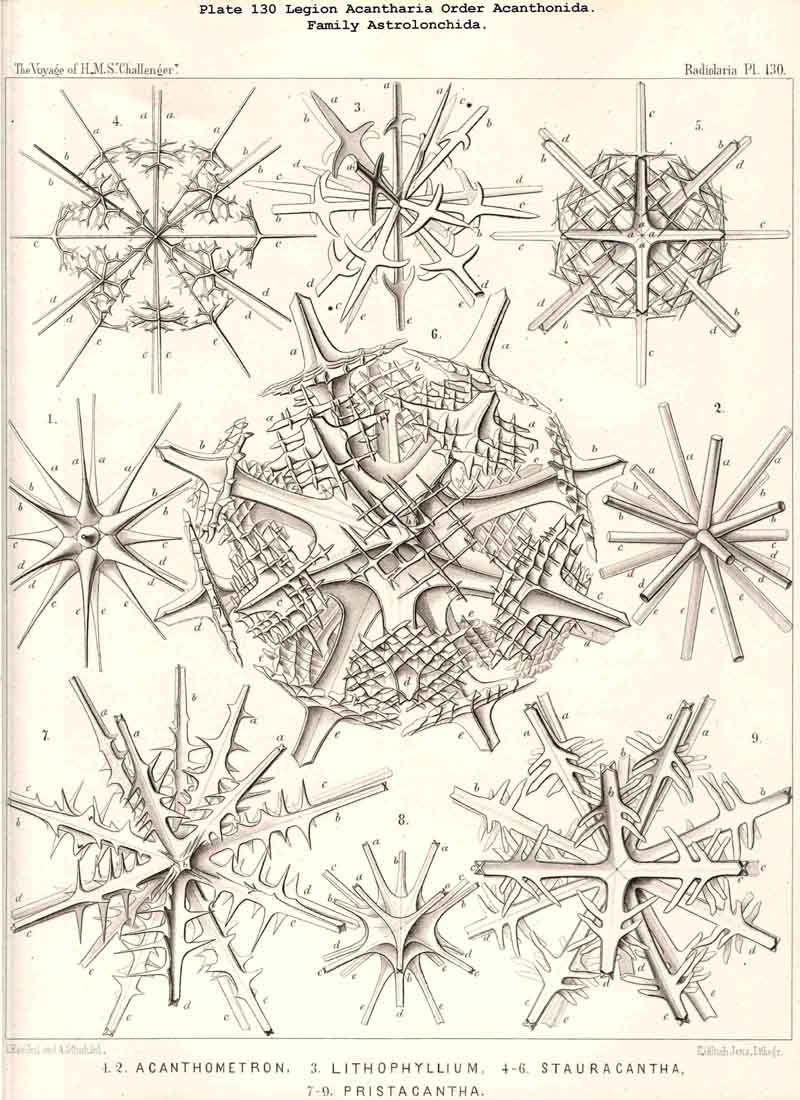{kind=link}
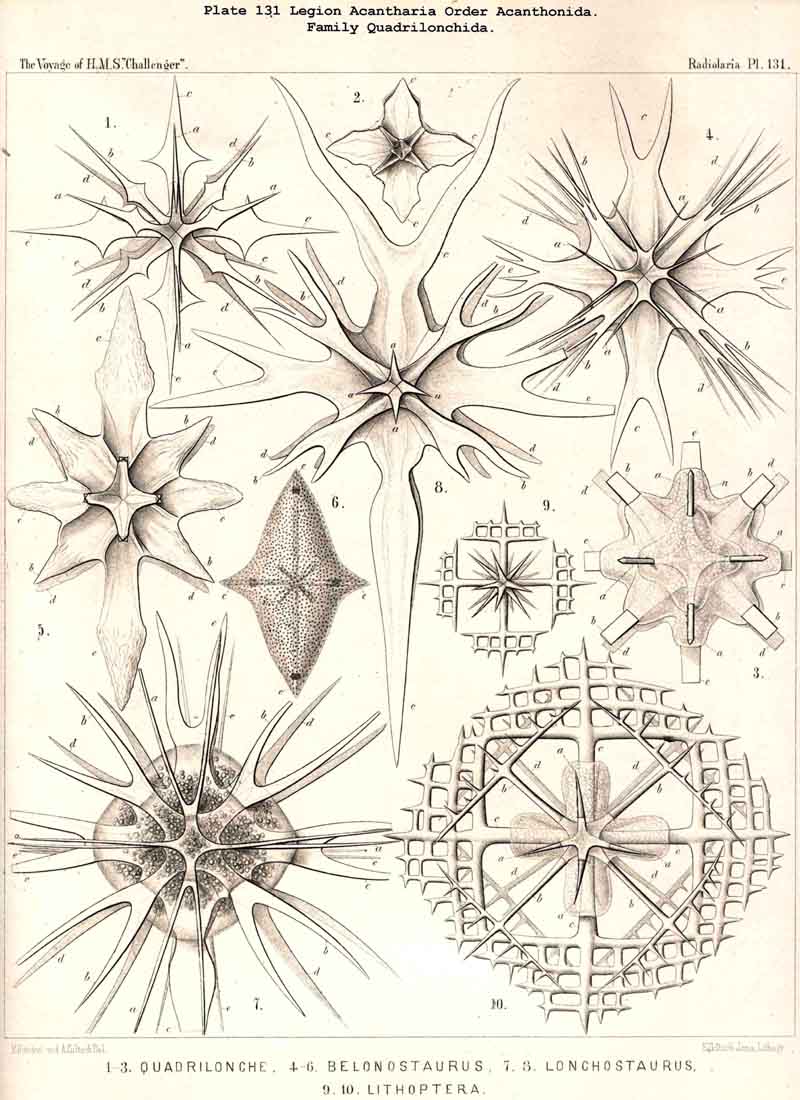{kind=link}
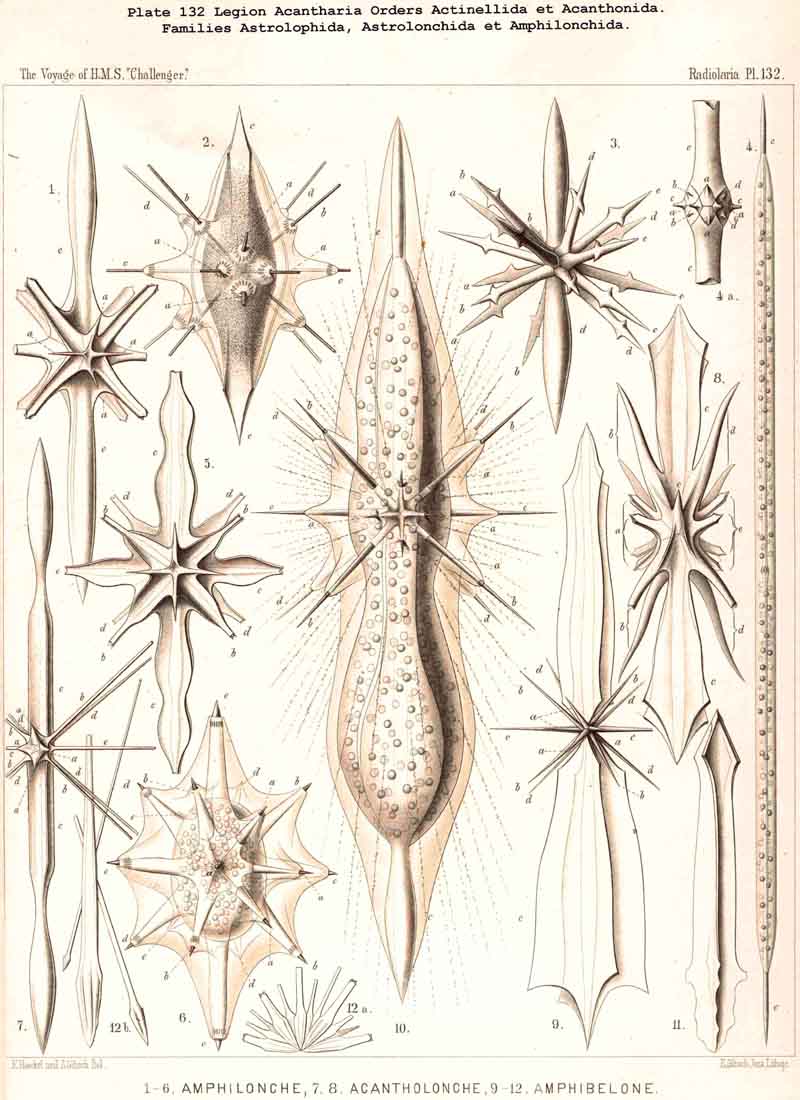{kind=link}
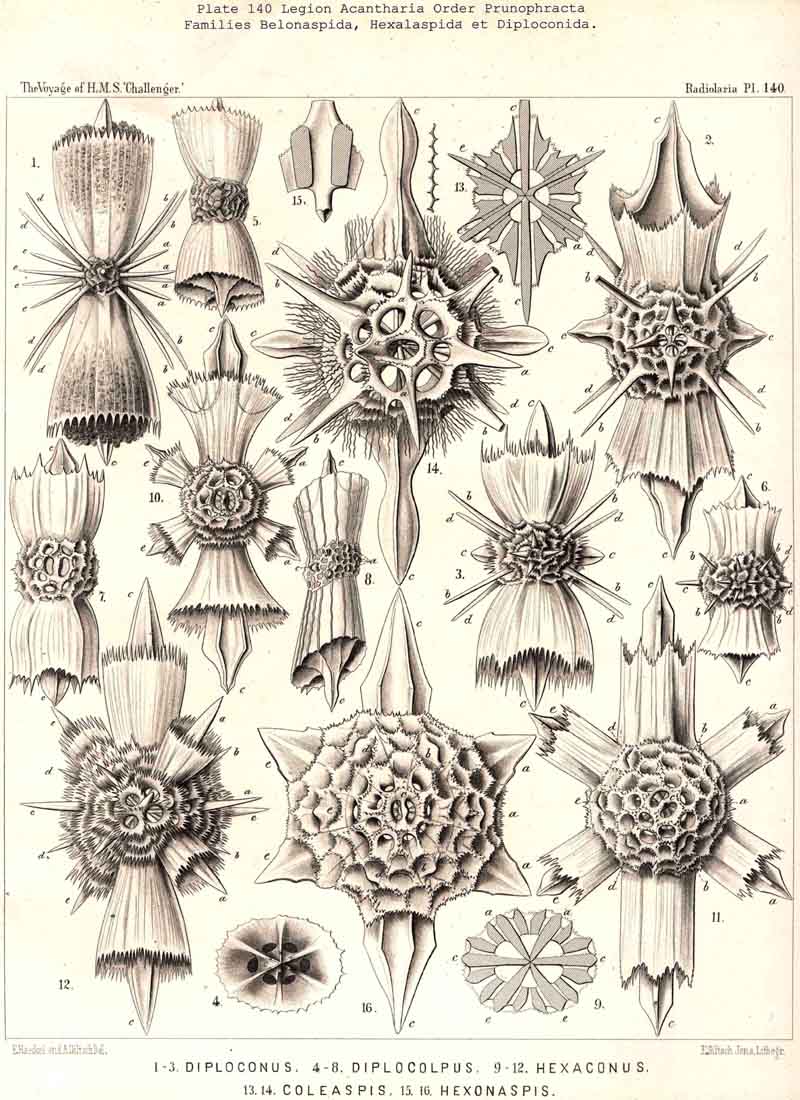{kind=link}
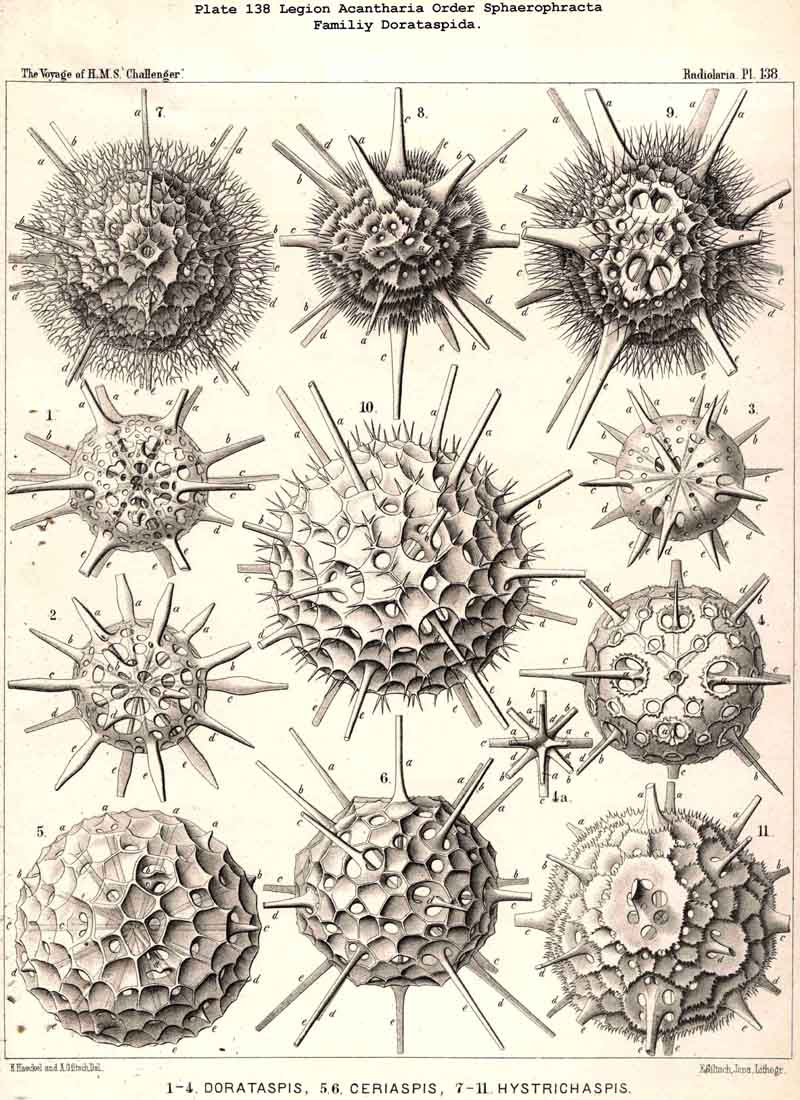{kind=link}
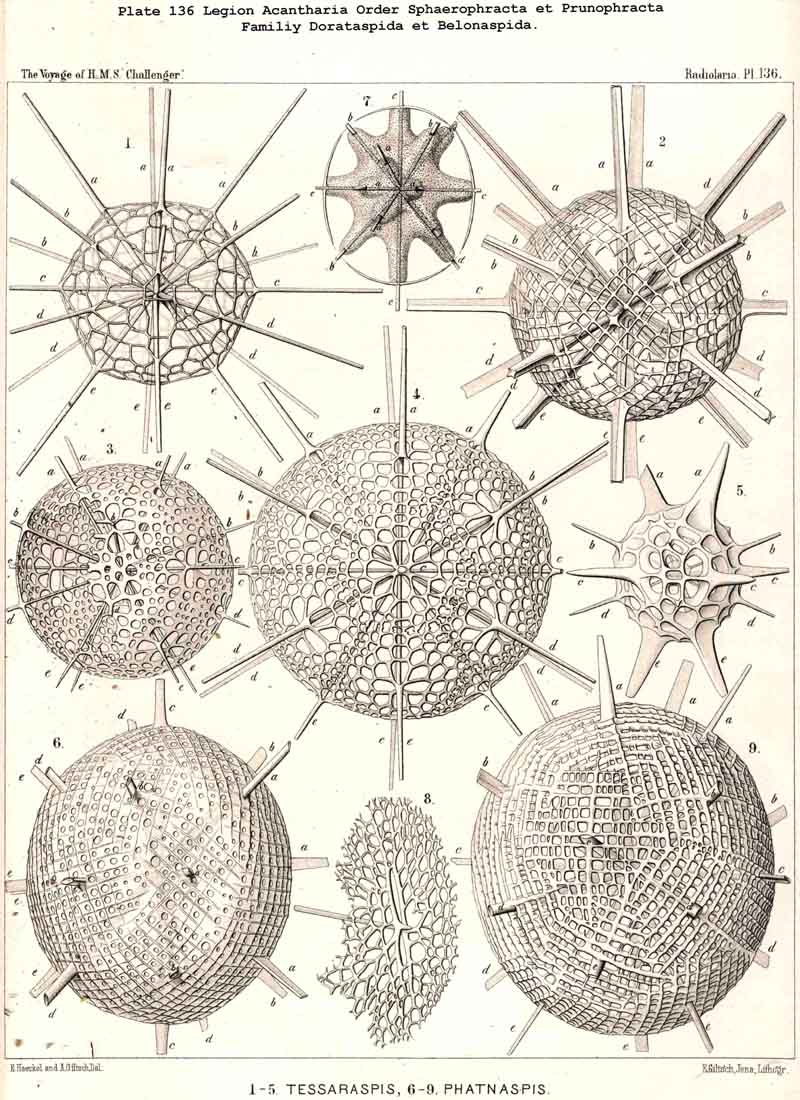{kind=link}
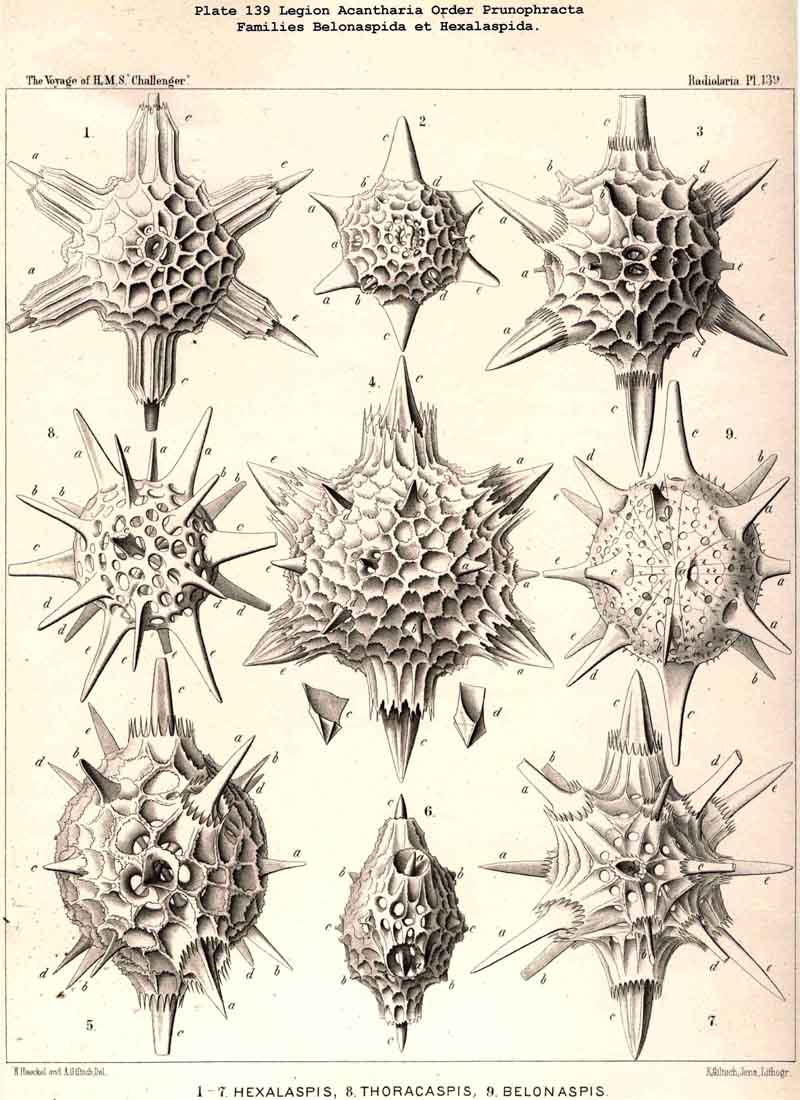{kind=link}
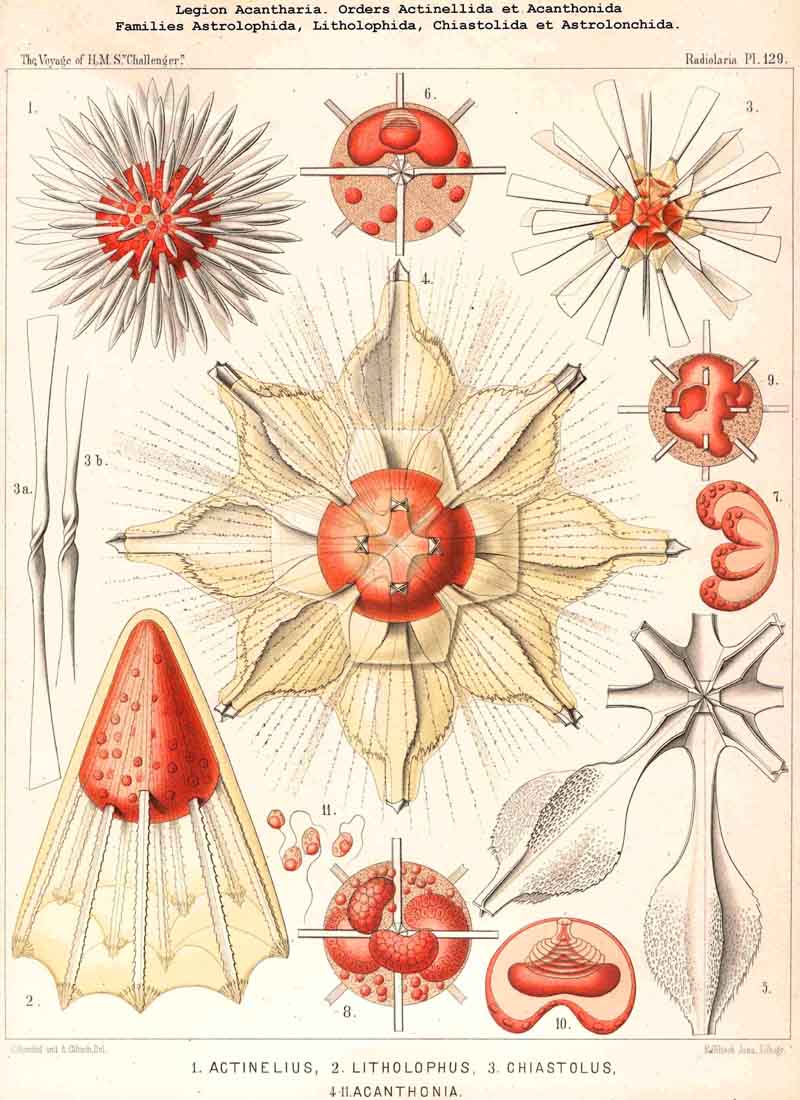{kind=link}
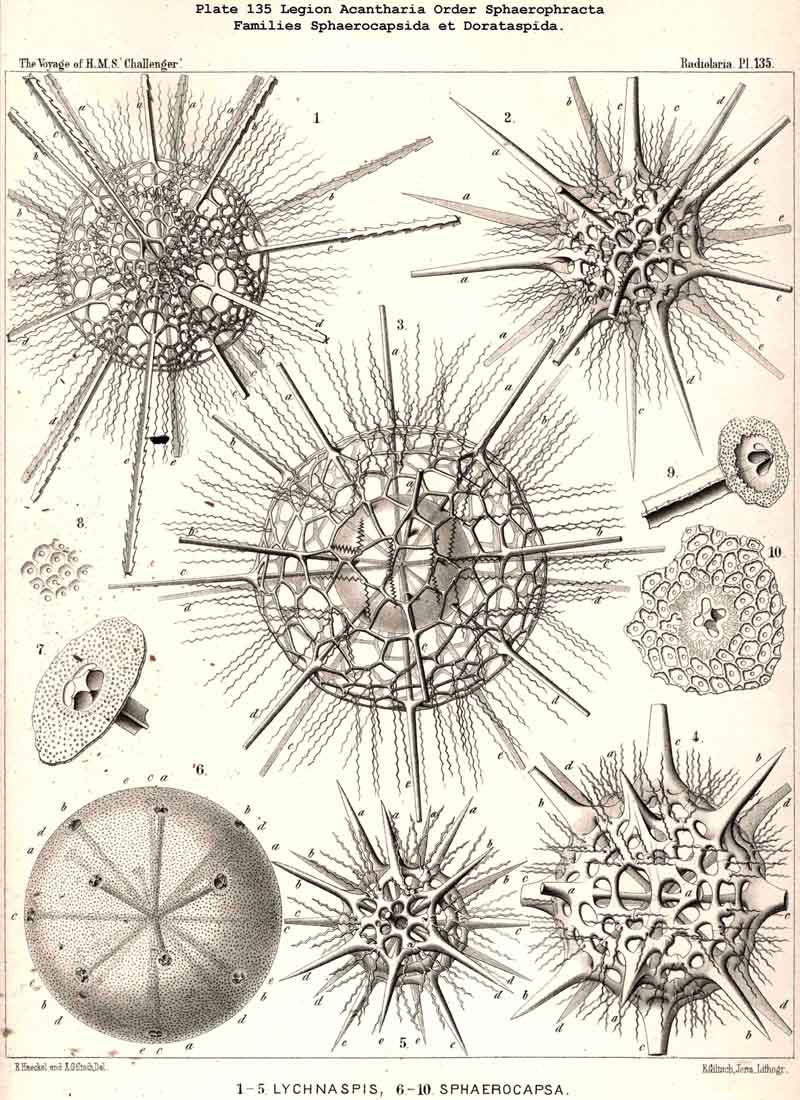{kind=link}
{kind=link}
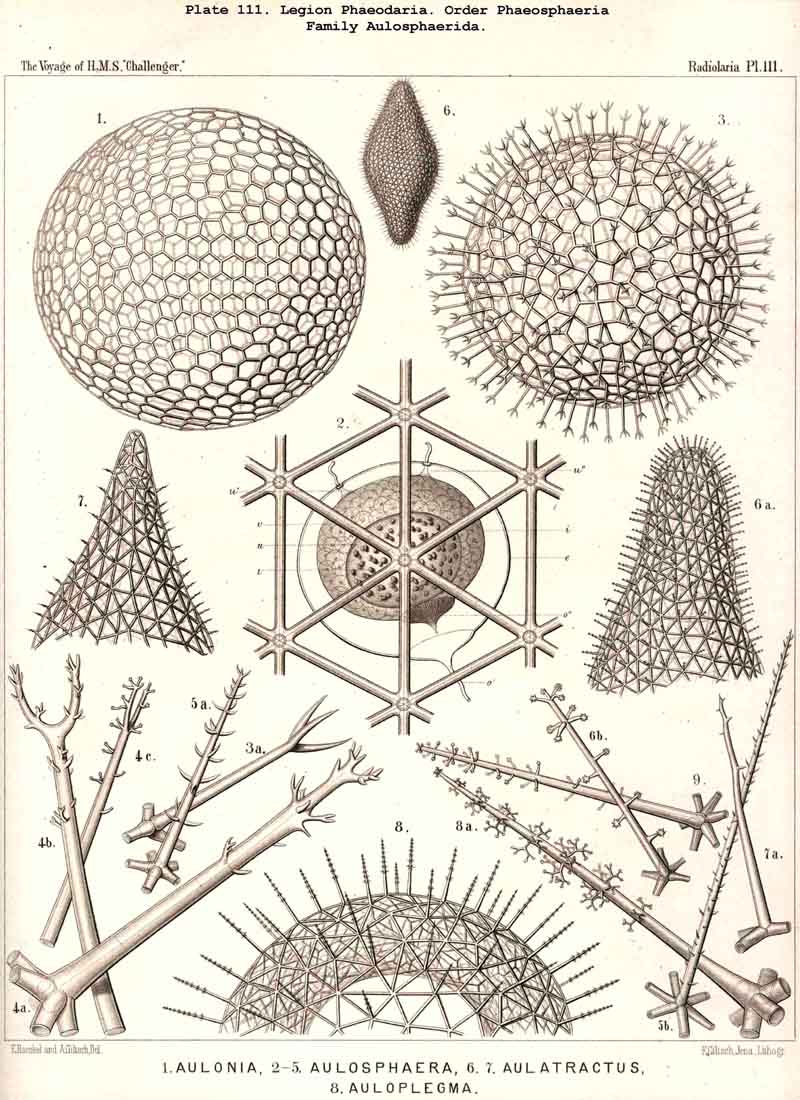{kind=link}
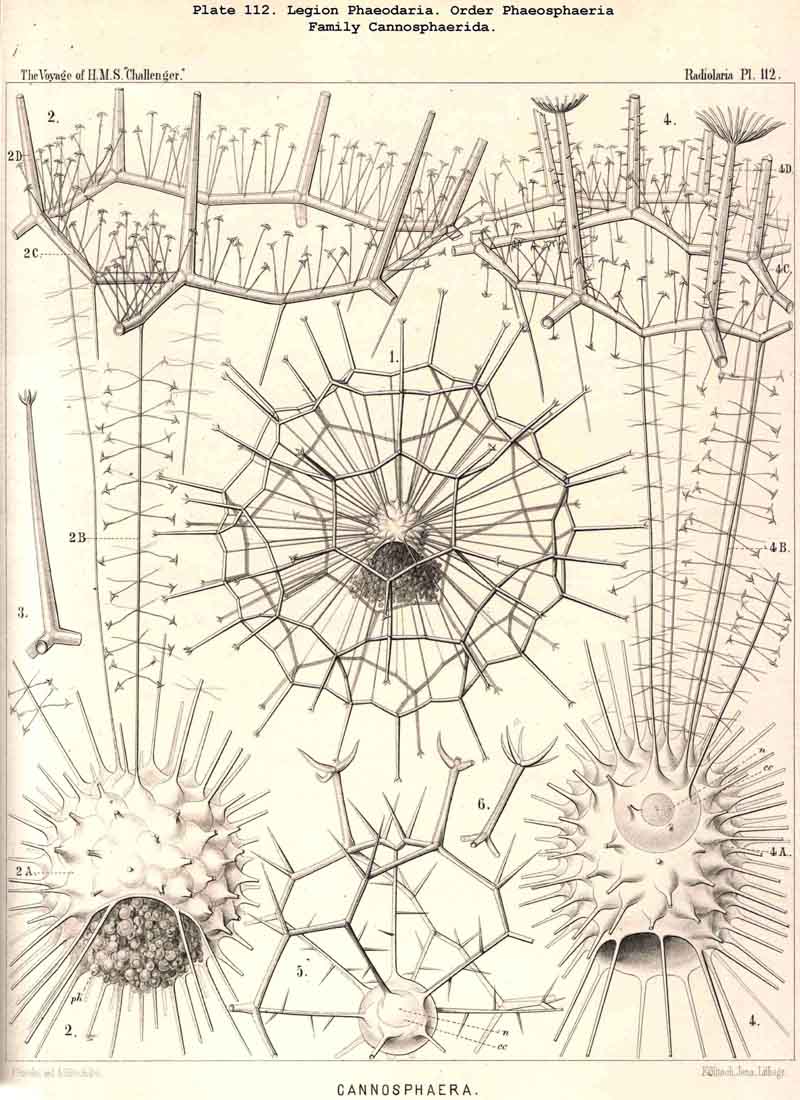{kind=link}
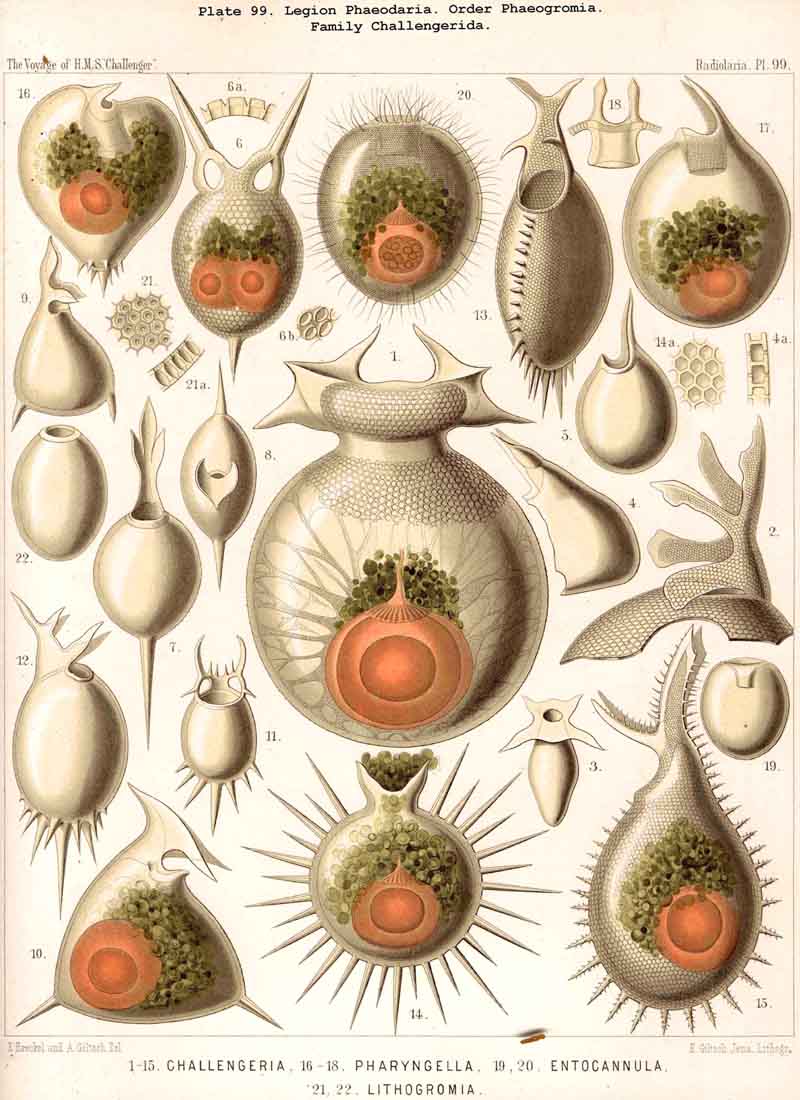{kind=link}
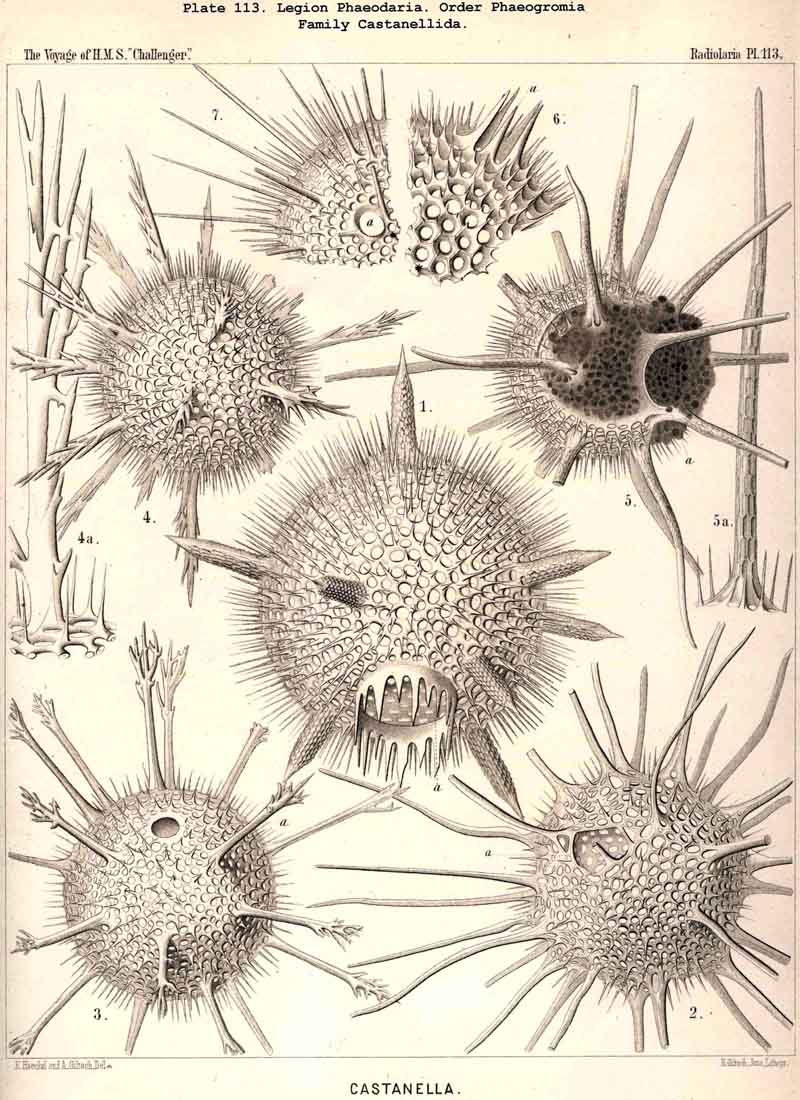{kind=link}
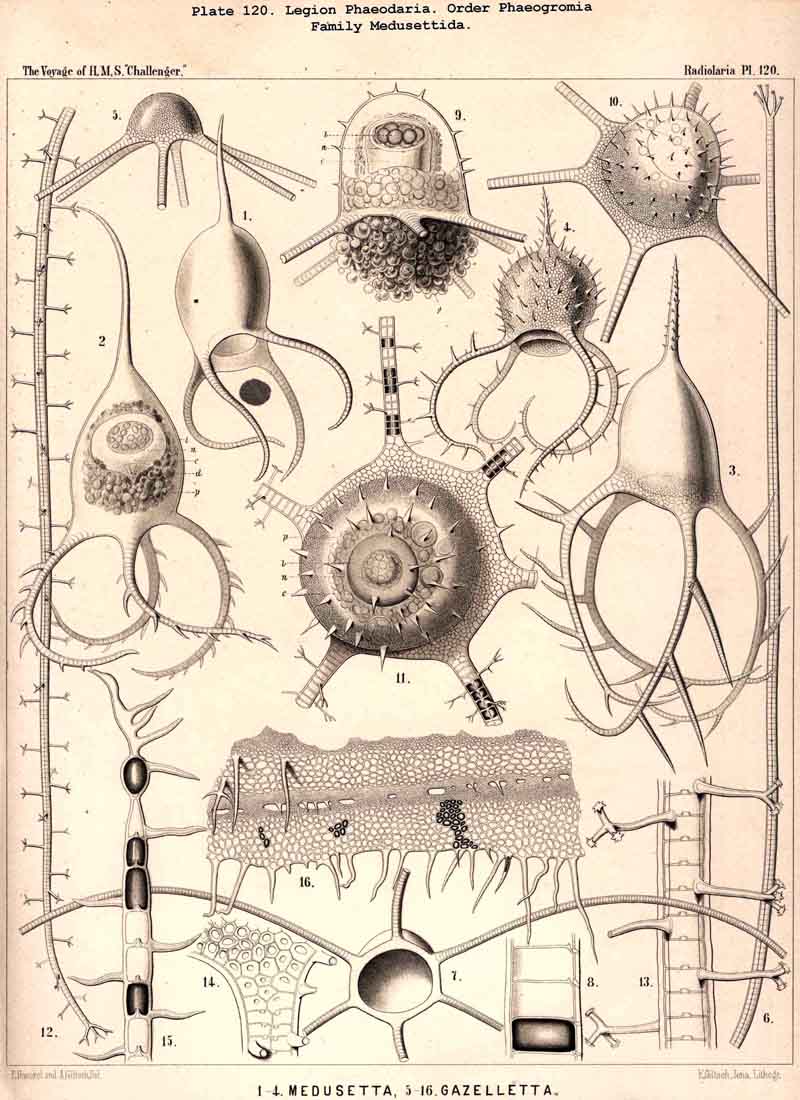{kind=link}
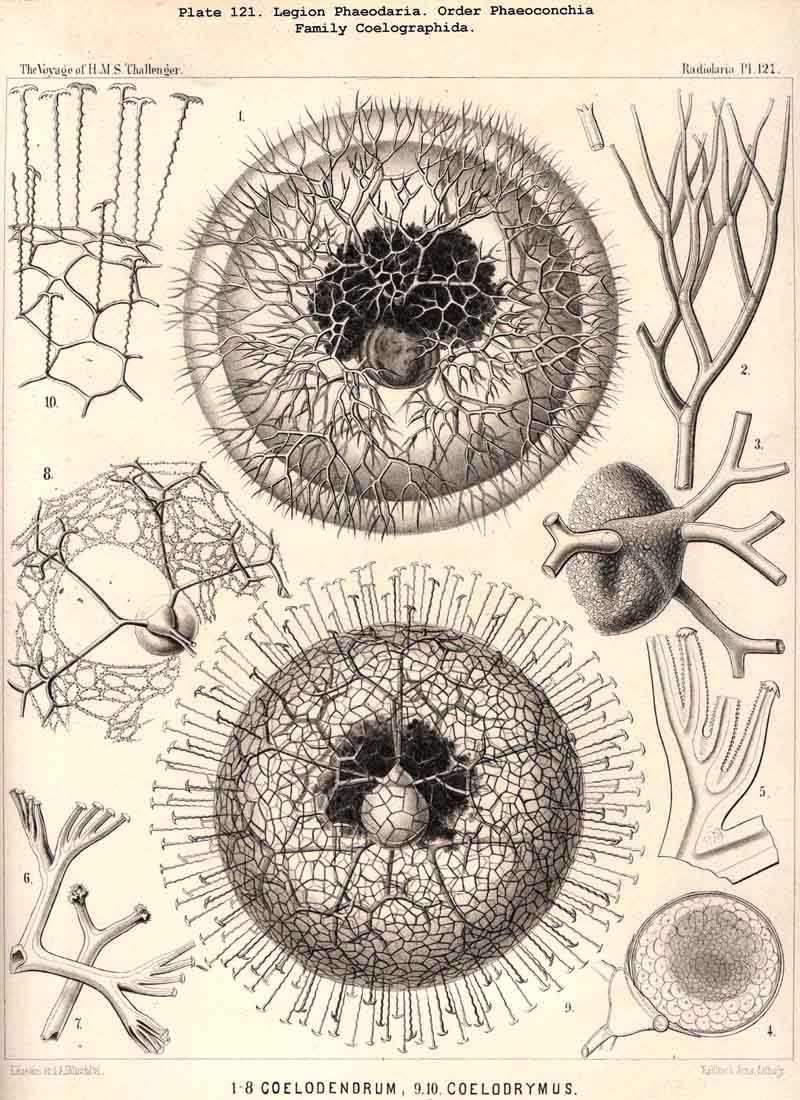{kind=link}
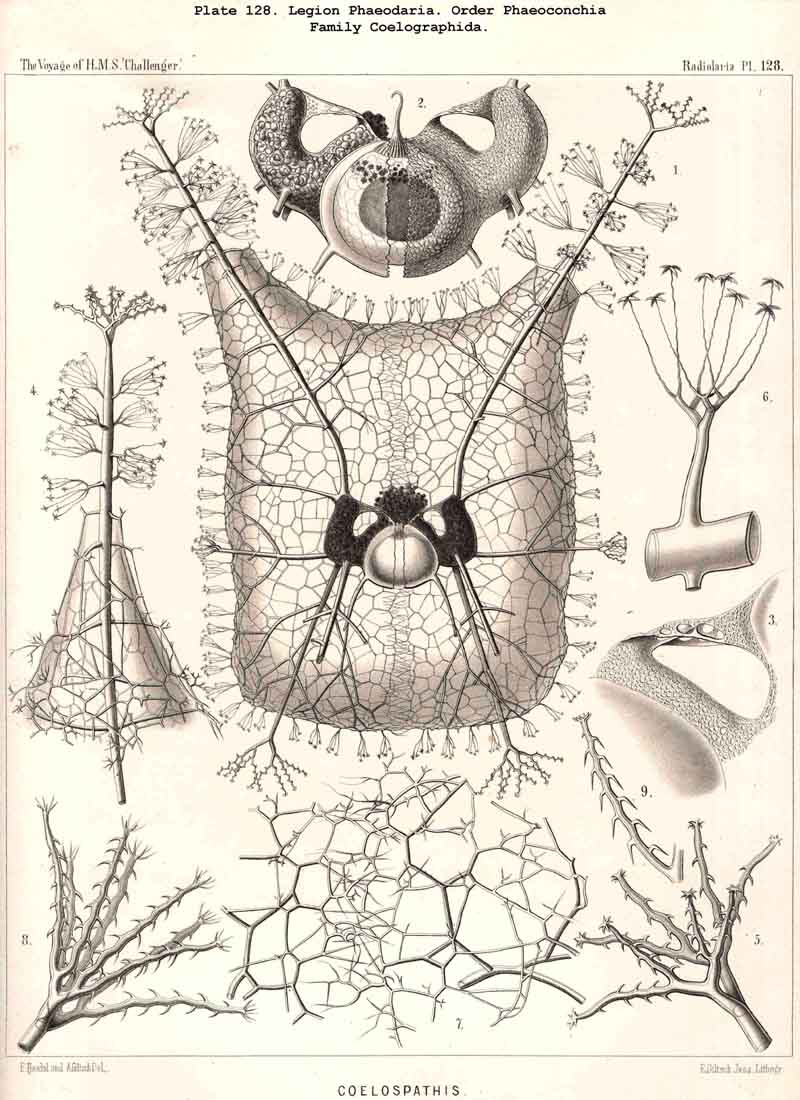{kind=link}
{kind=link}
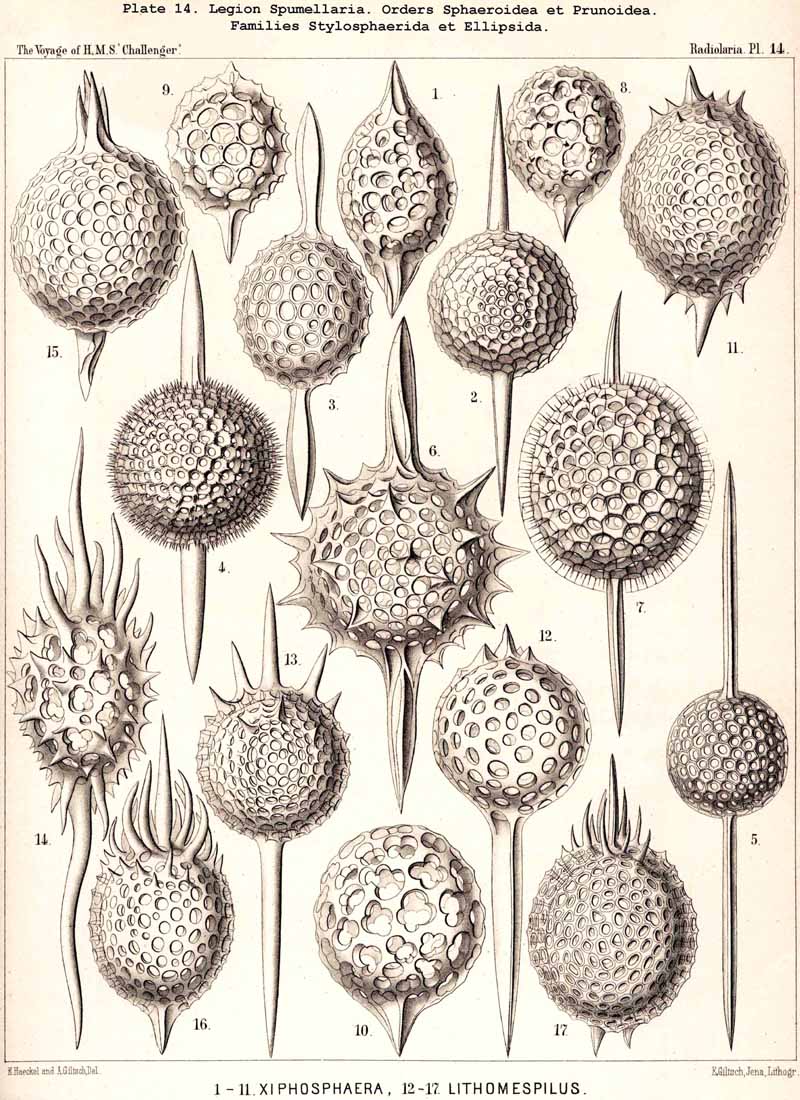{kind=link}
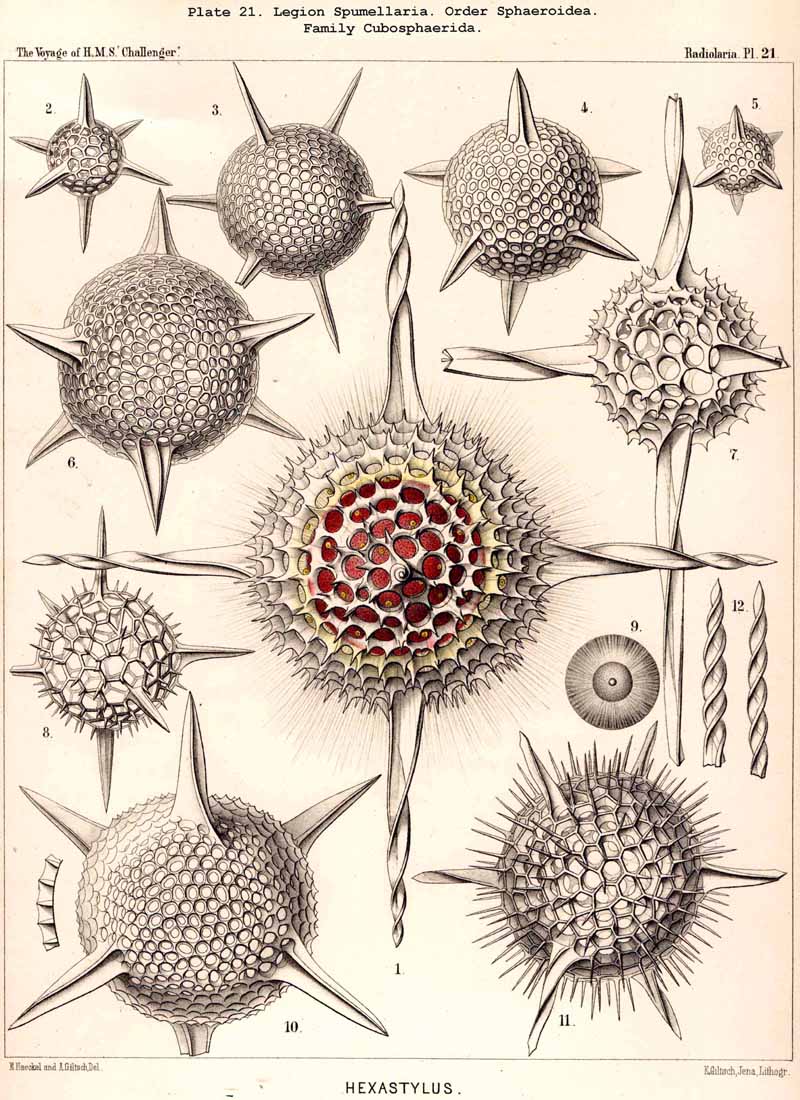{kind=link}
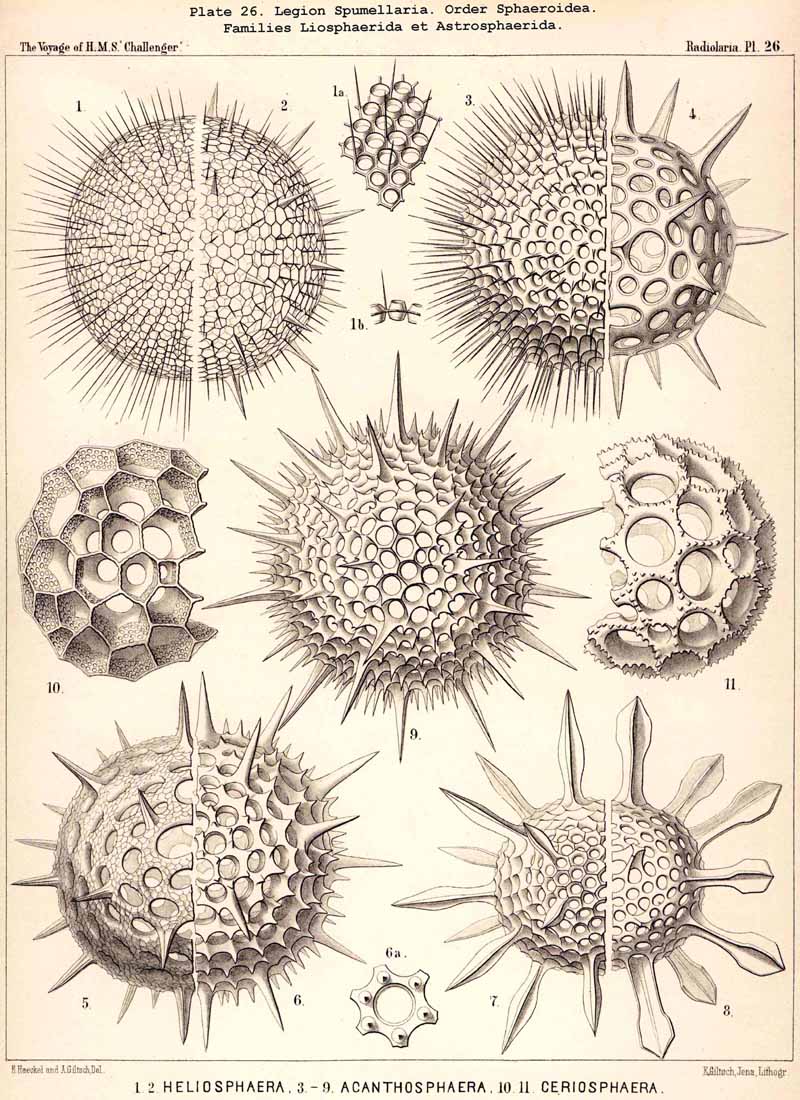{kind=link}
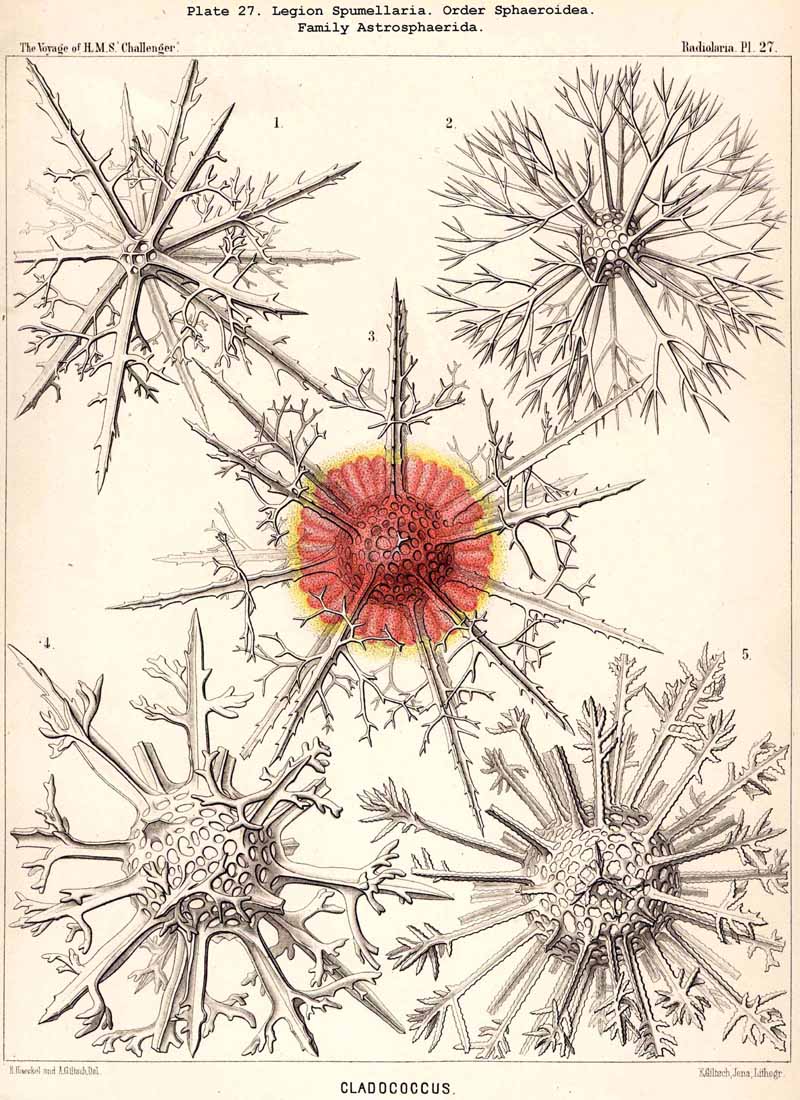{kind=link}
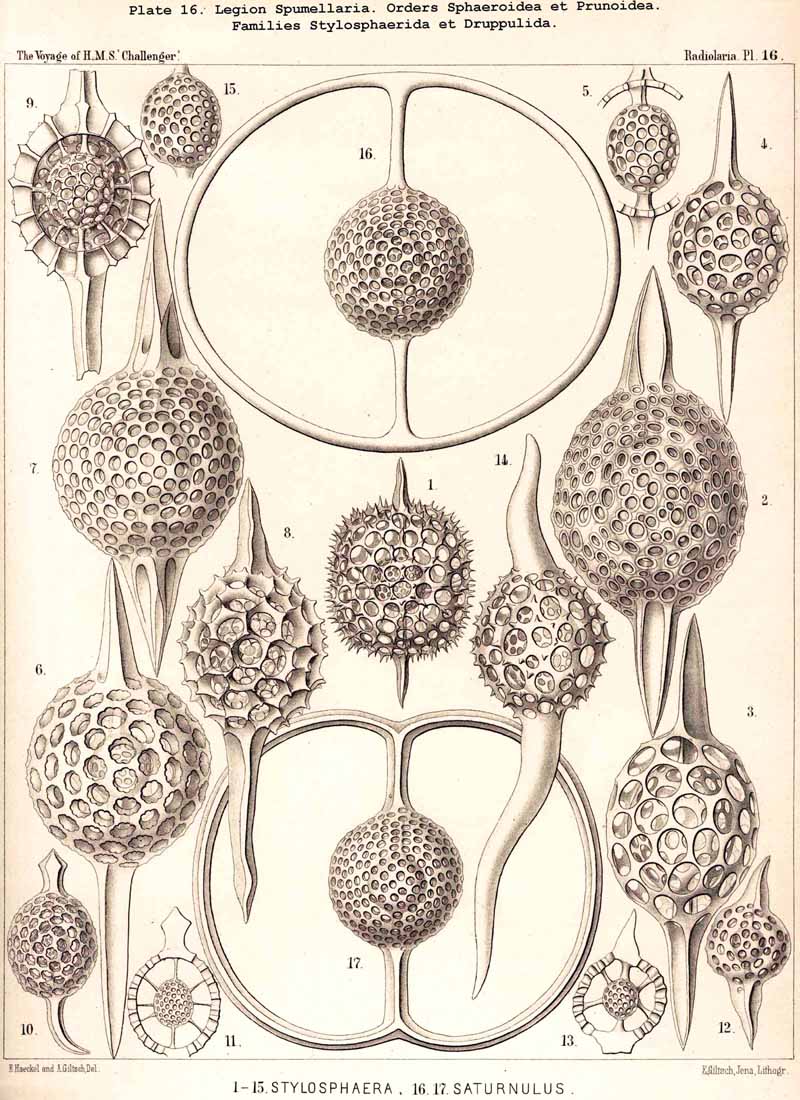{kind=link}
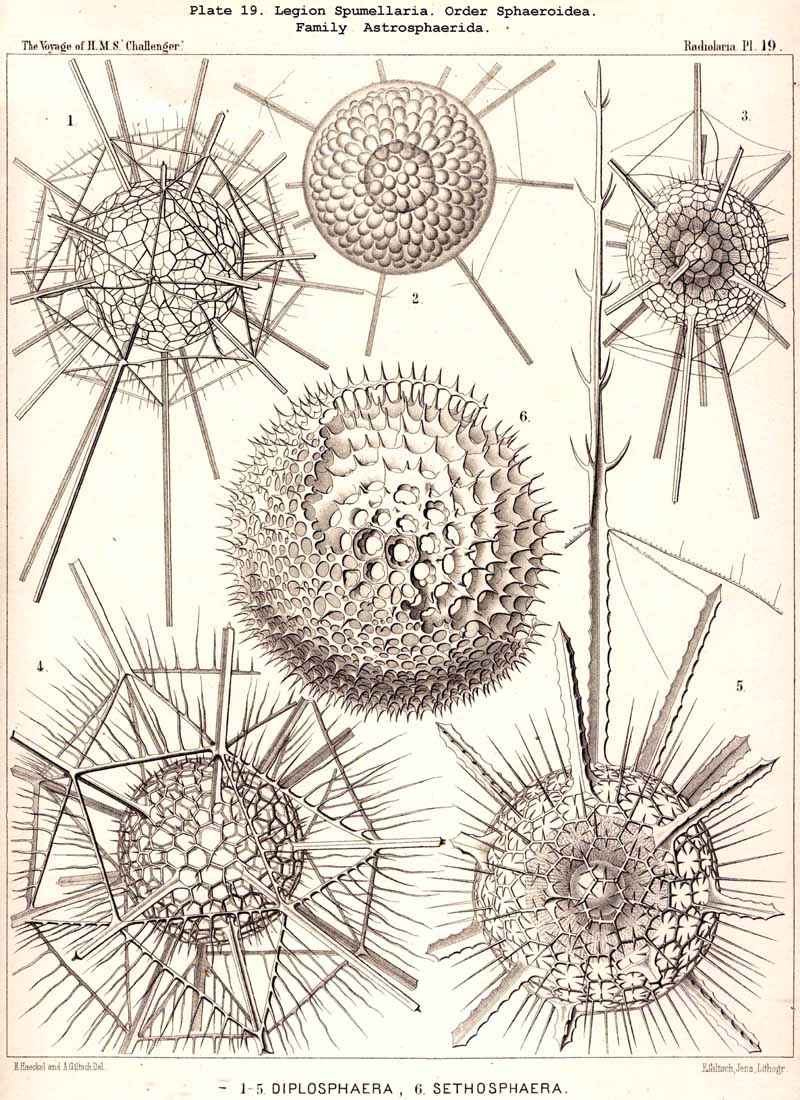{kind=link}
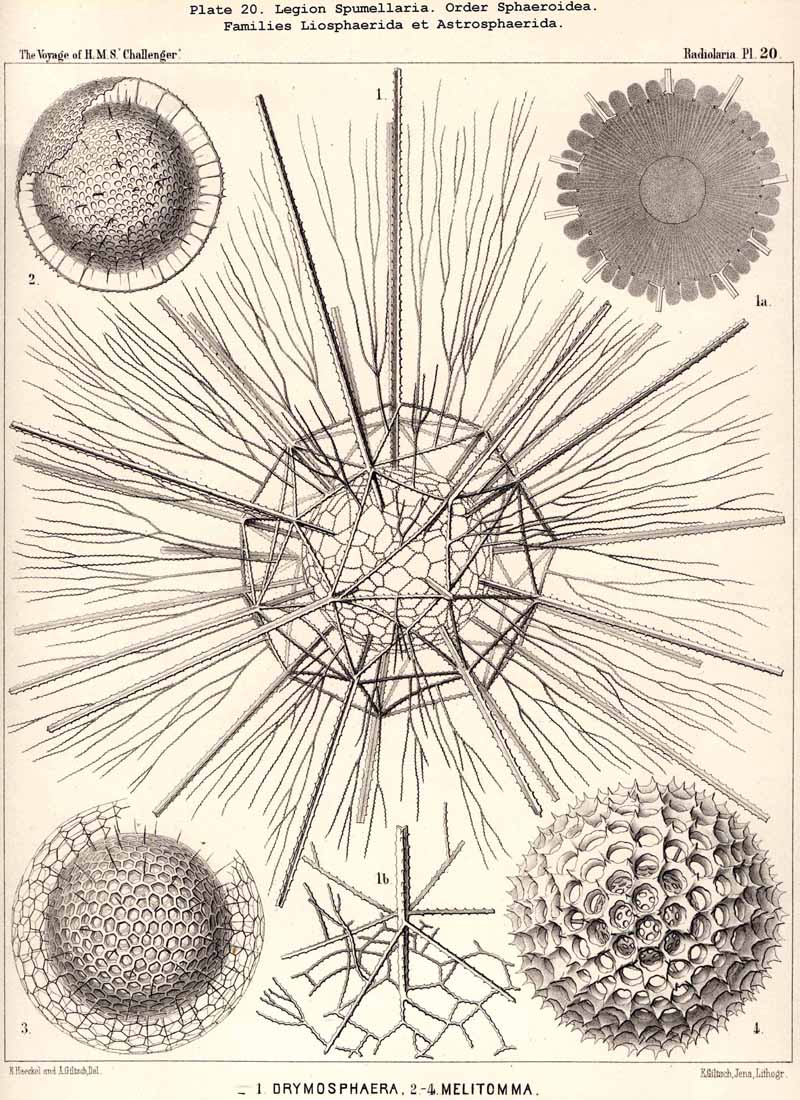{kind=link}
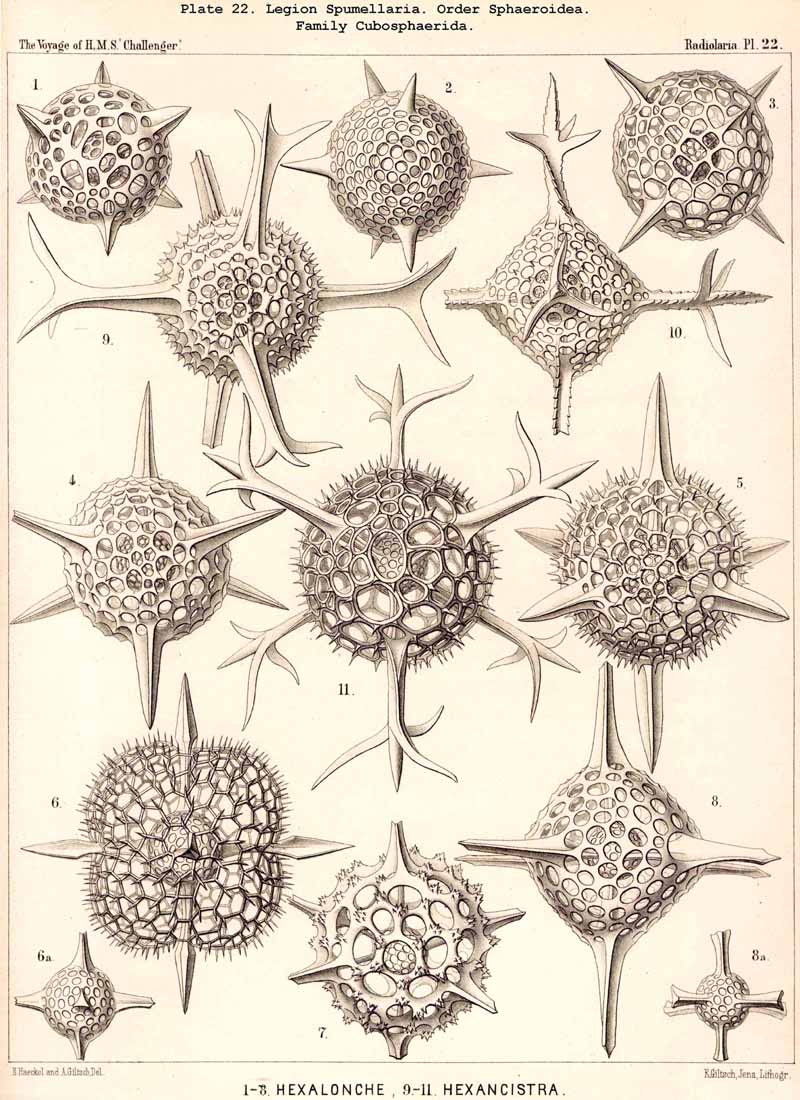{kind=link}
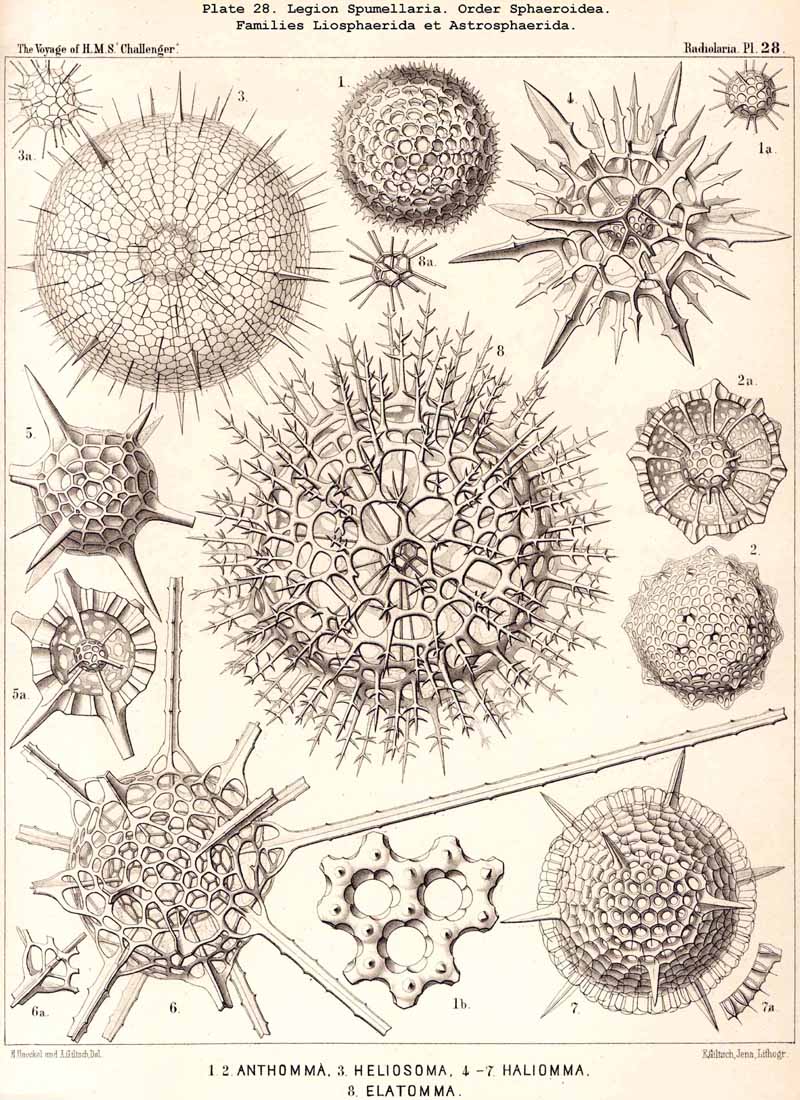{kind=link}
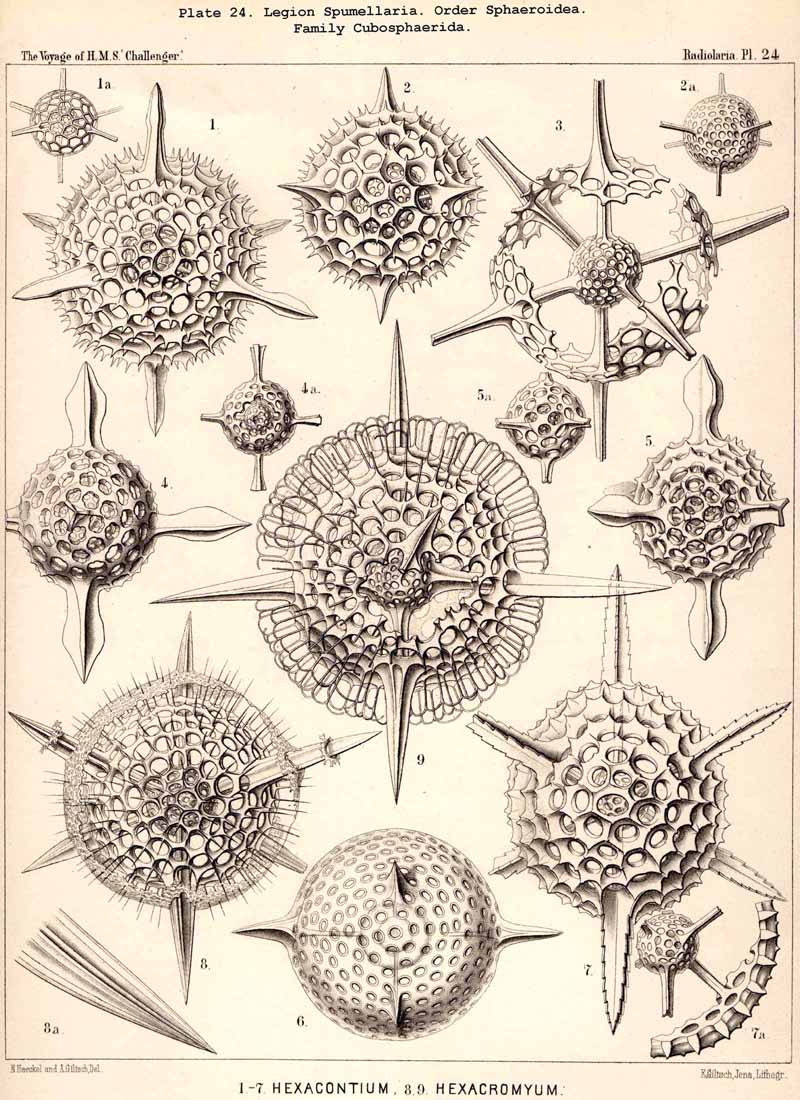{kind=link}
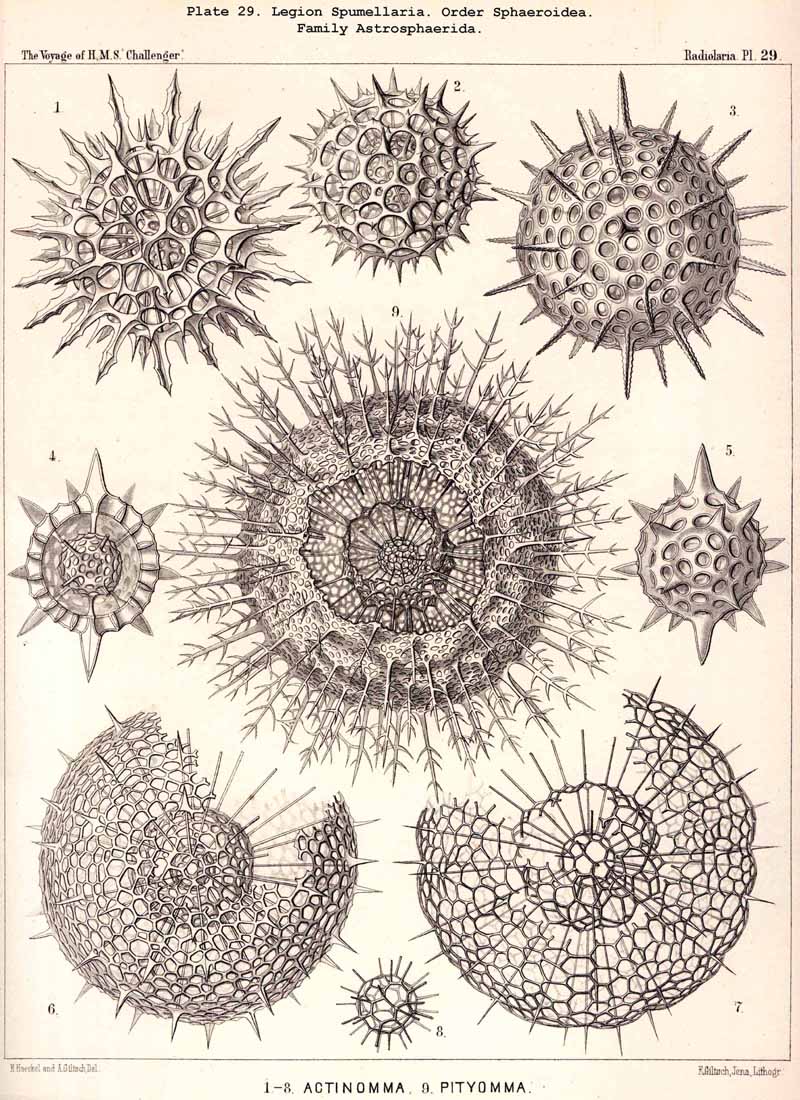{kind=link}
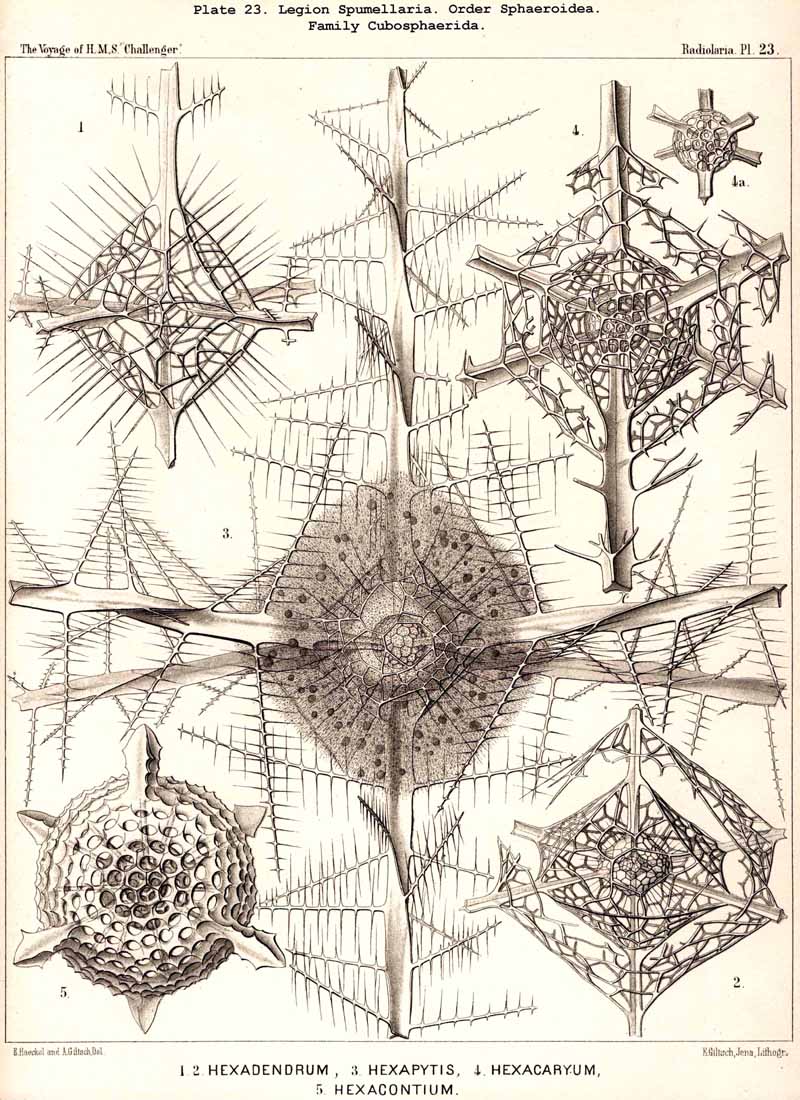{kind=link}
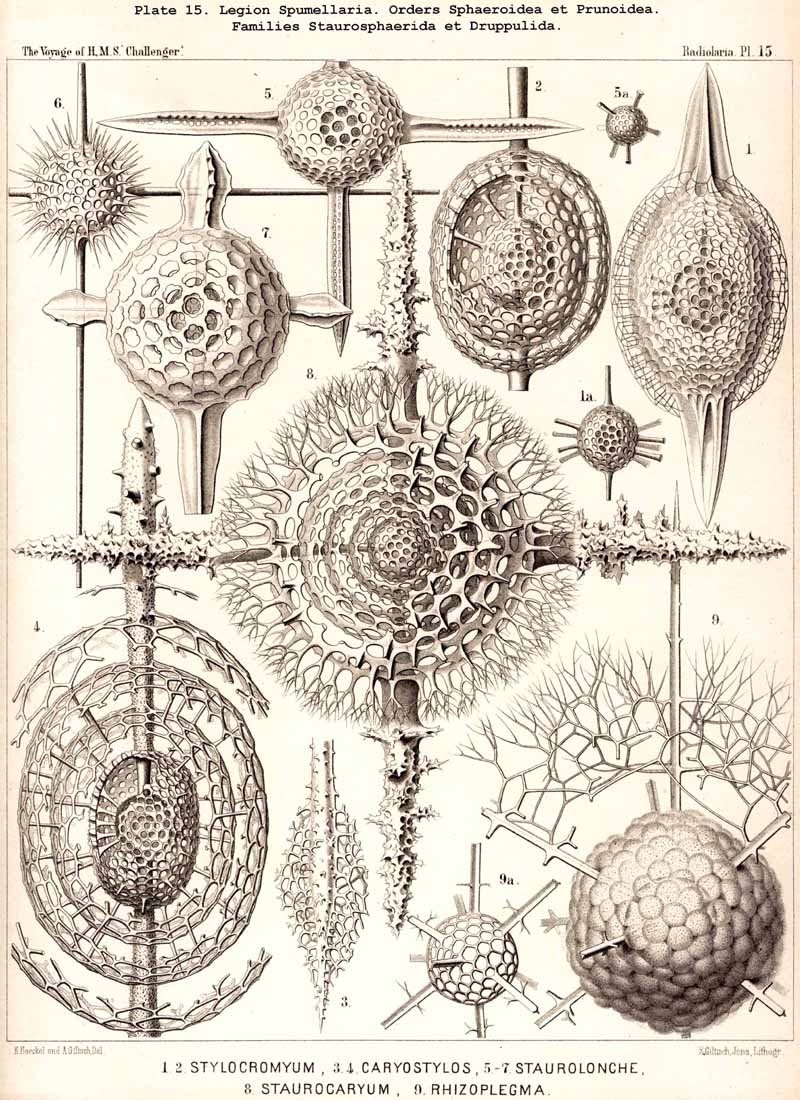{kind=link}
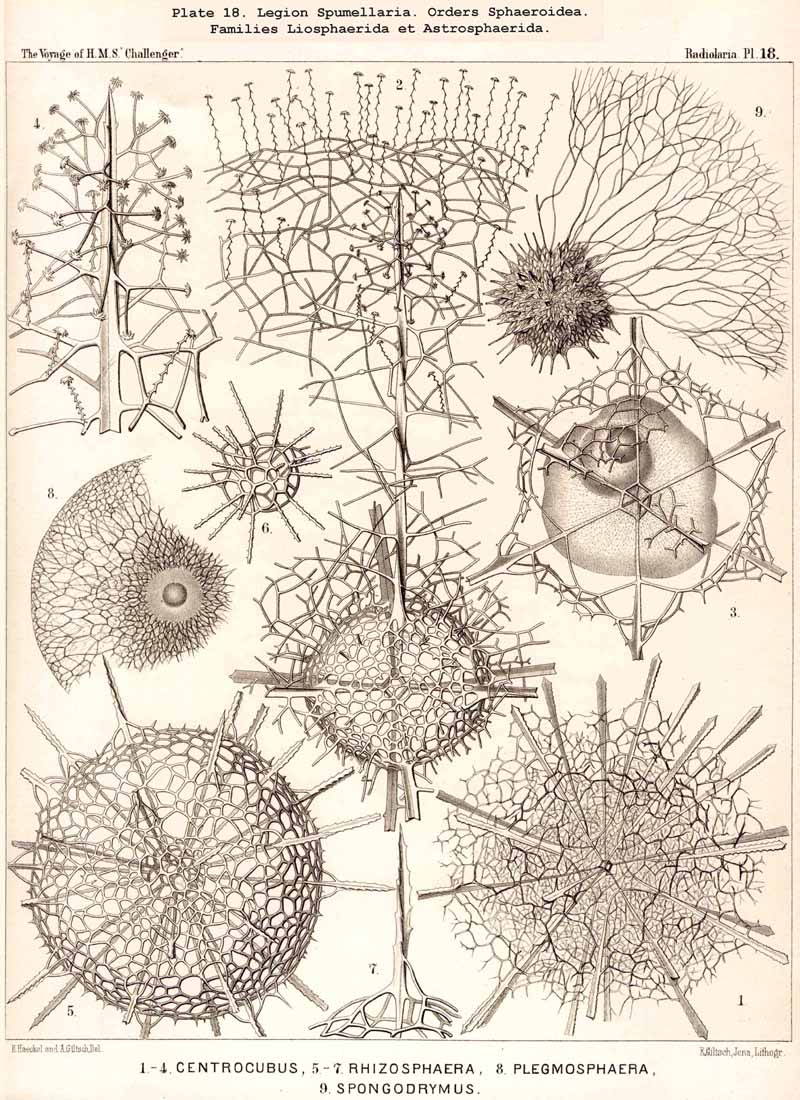{kind=link}
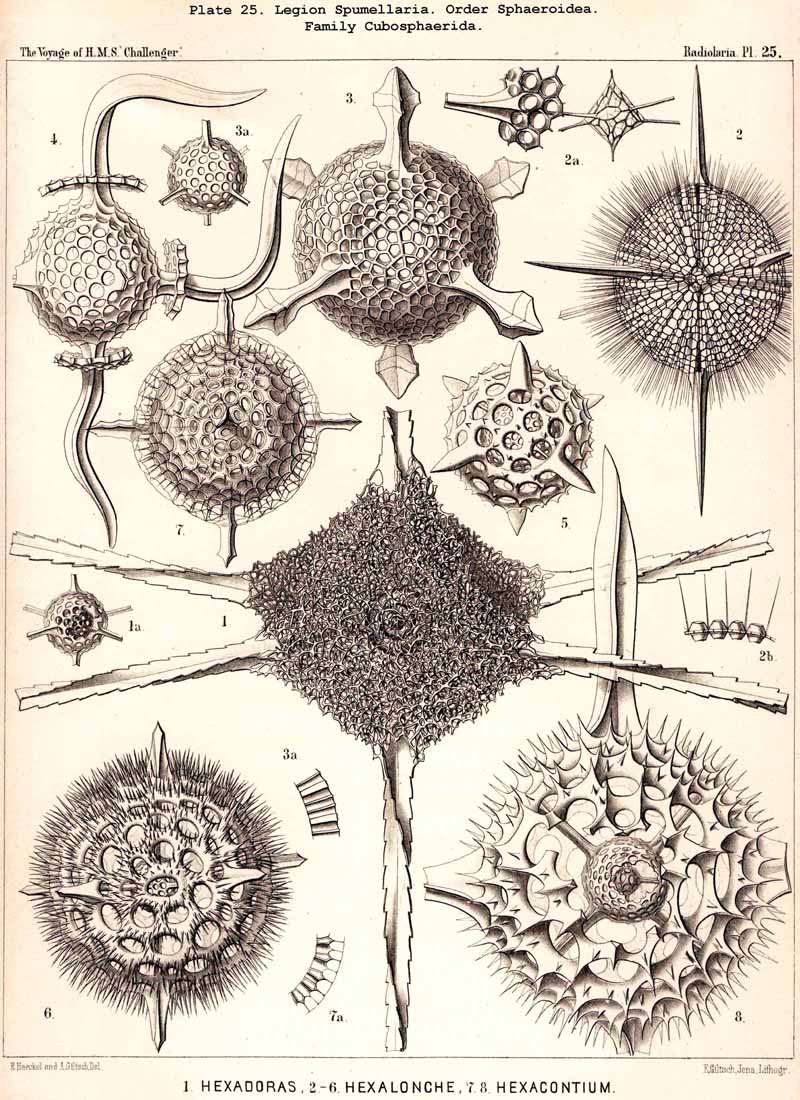{kind=link}
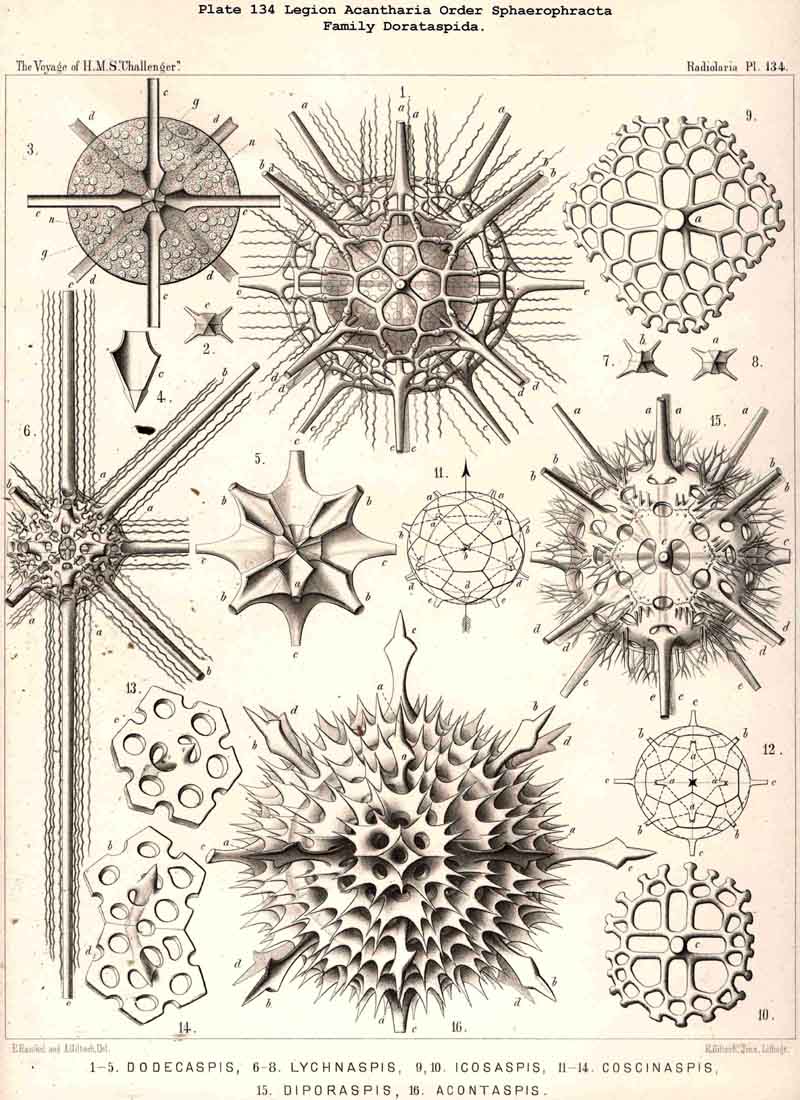{kind=link}
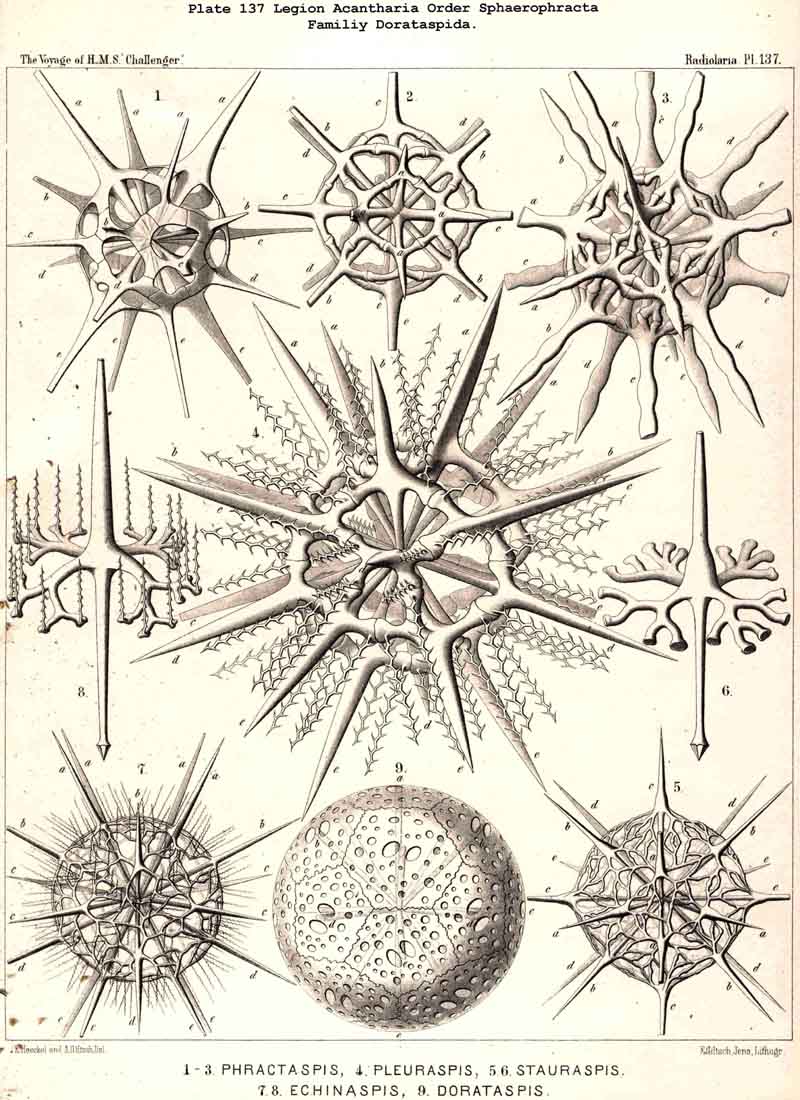{kind=link}
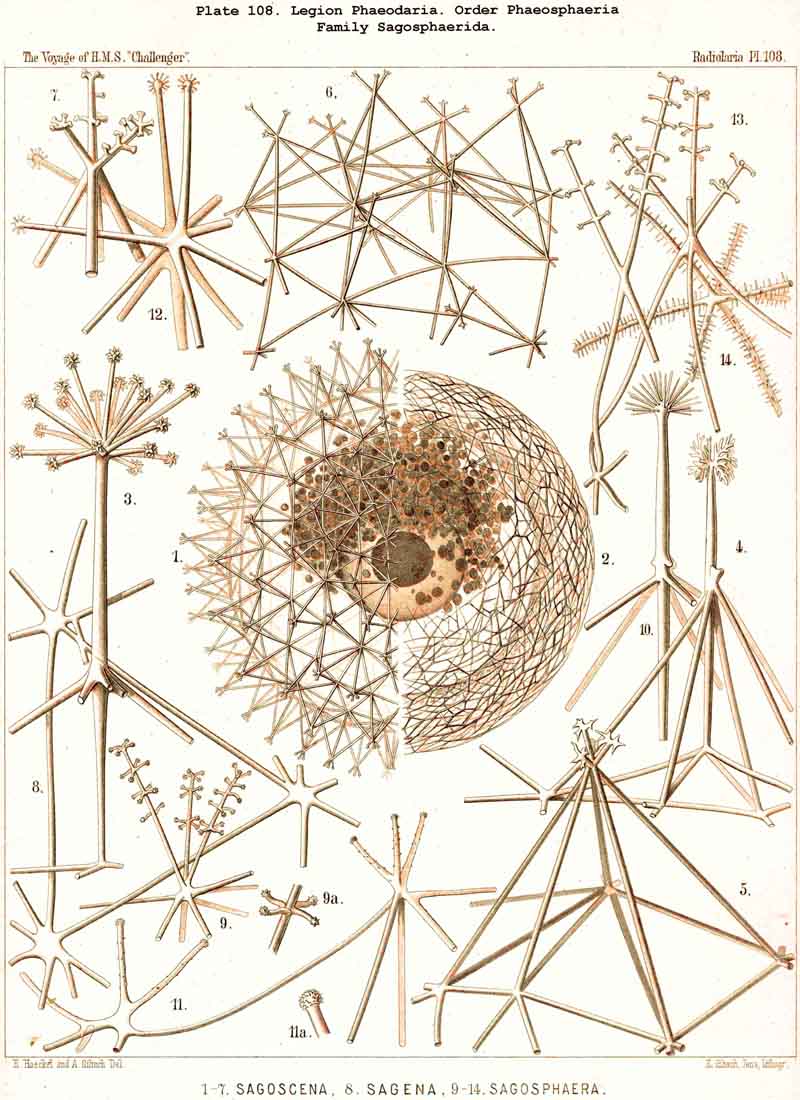{kind=link}
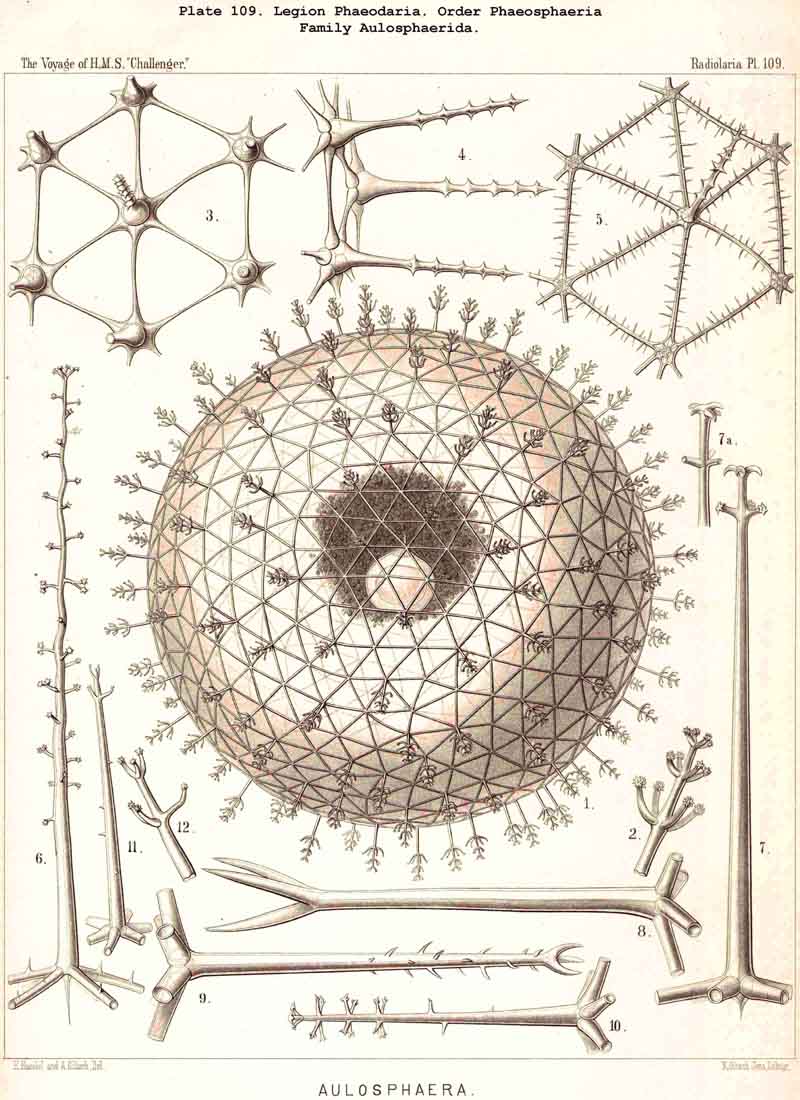{kind=link}
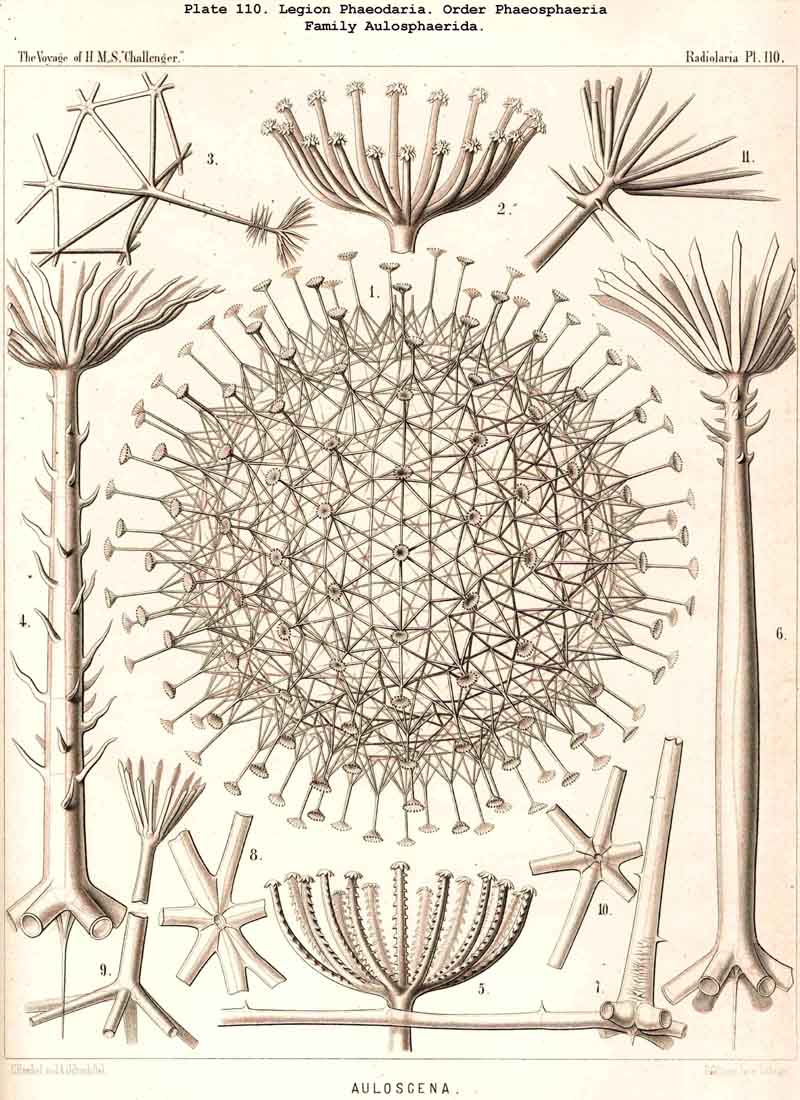{kind=link}
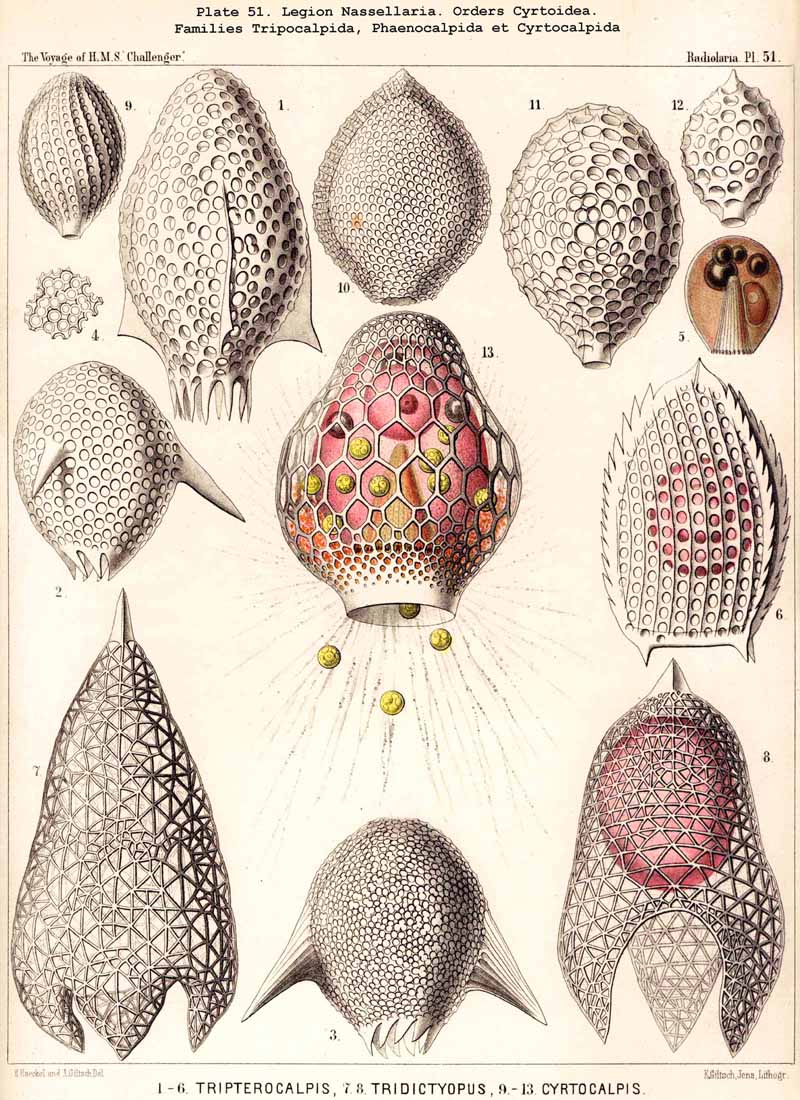{kind=link}
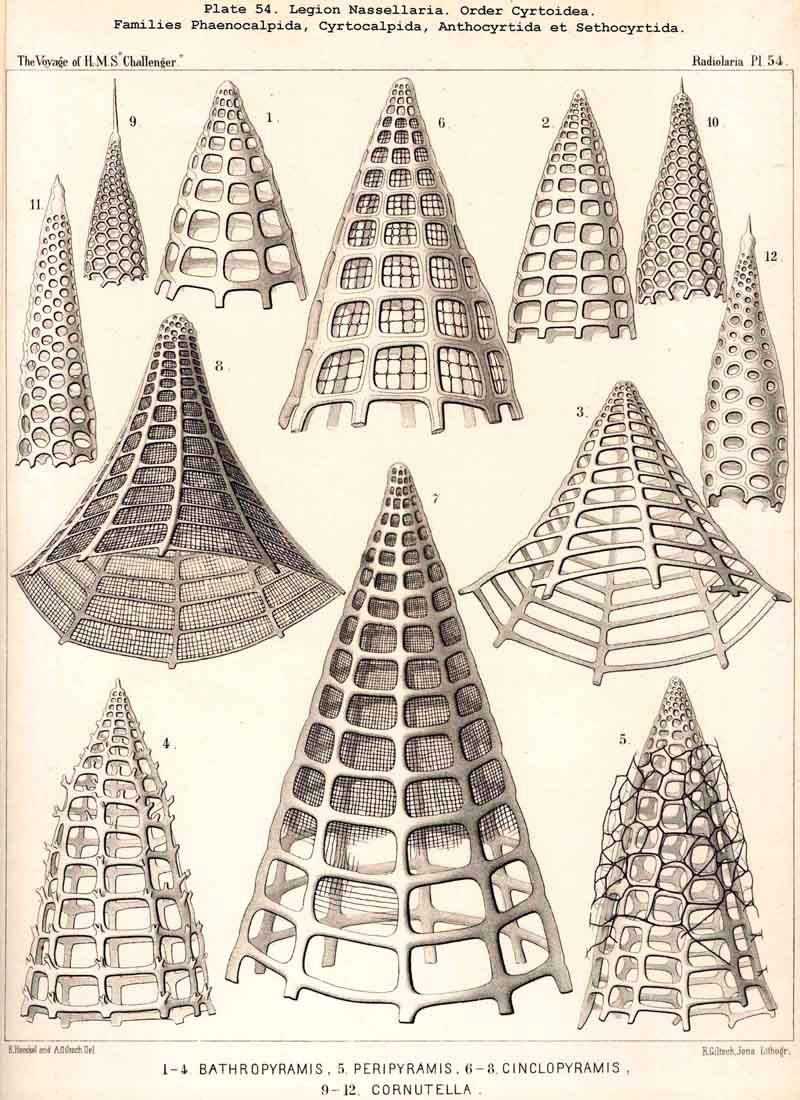{kind=link}
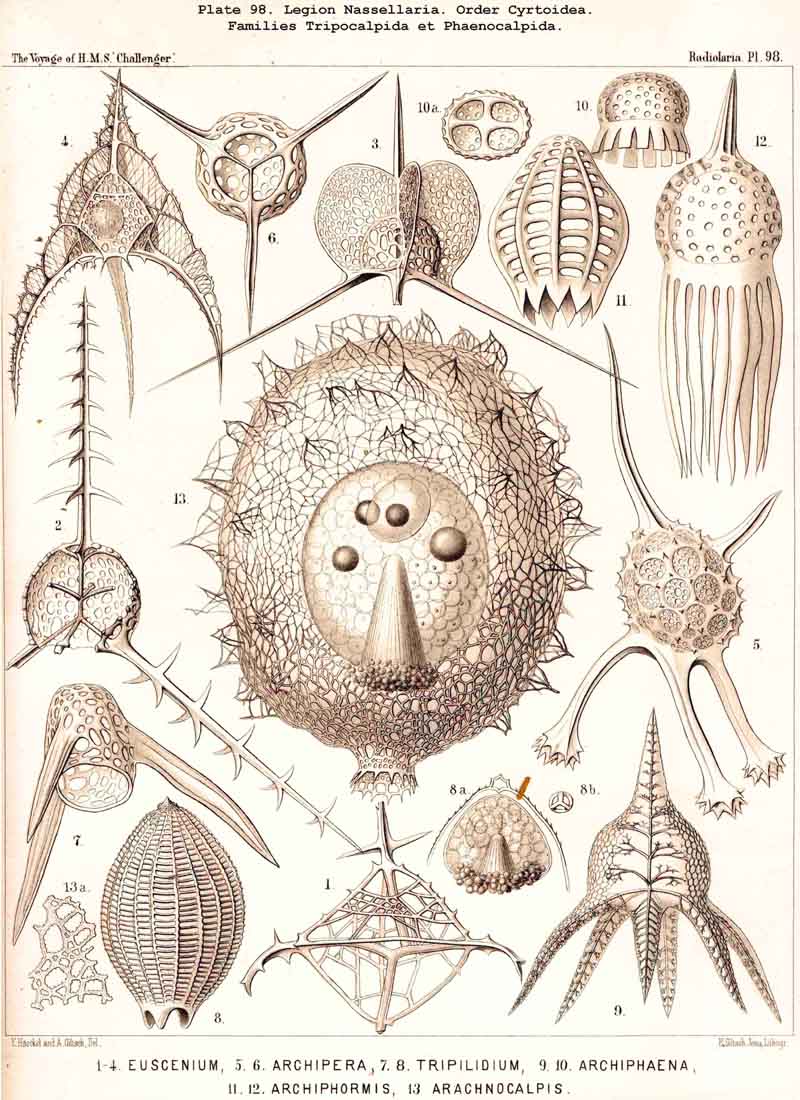{kind=link}
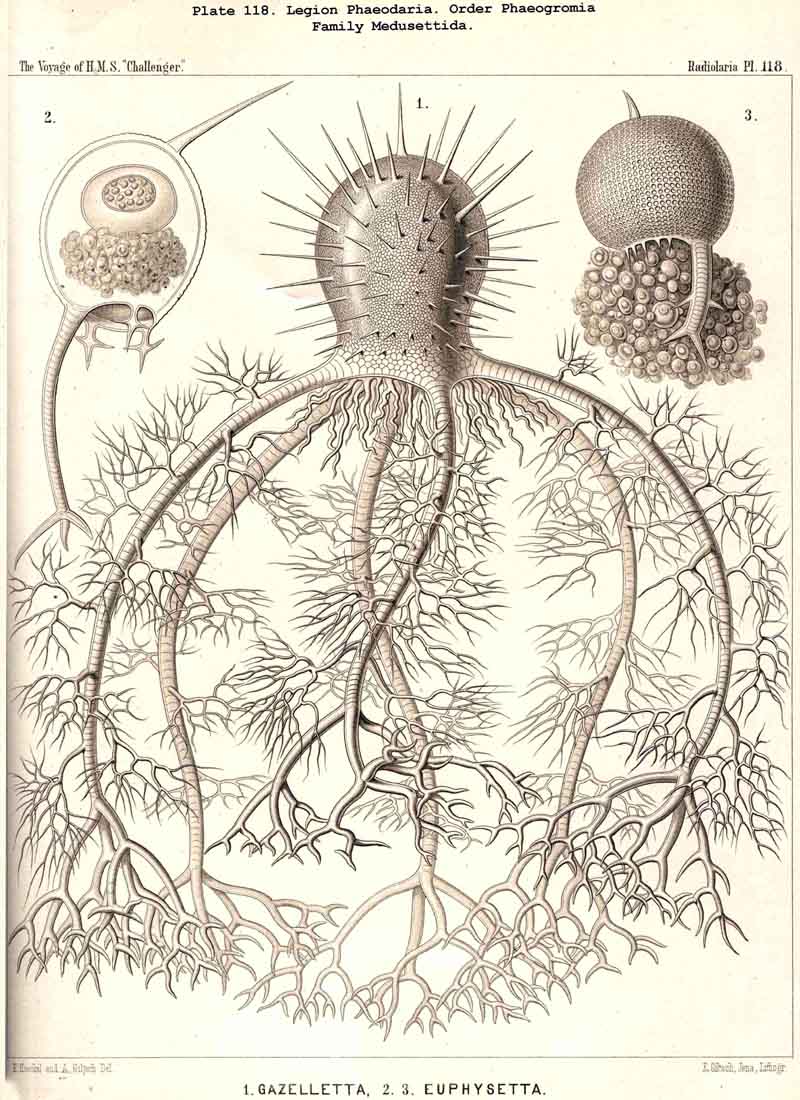{kind=link}
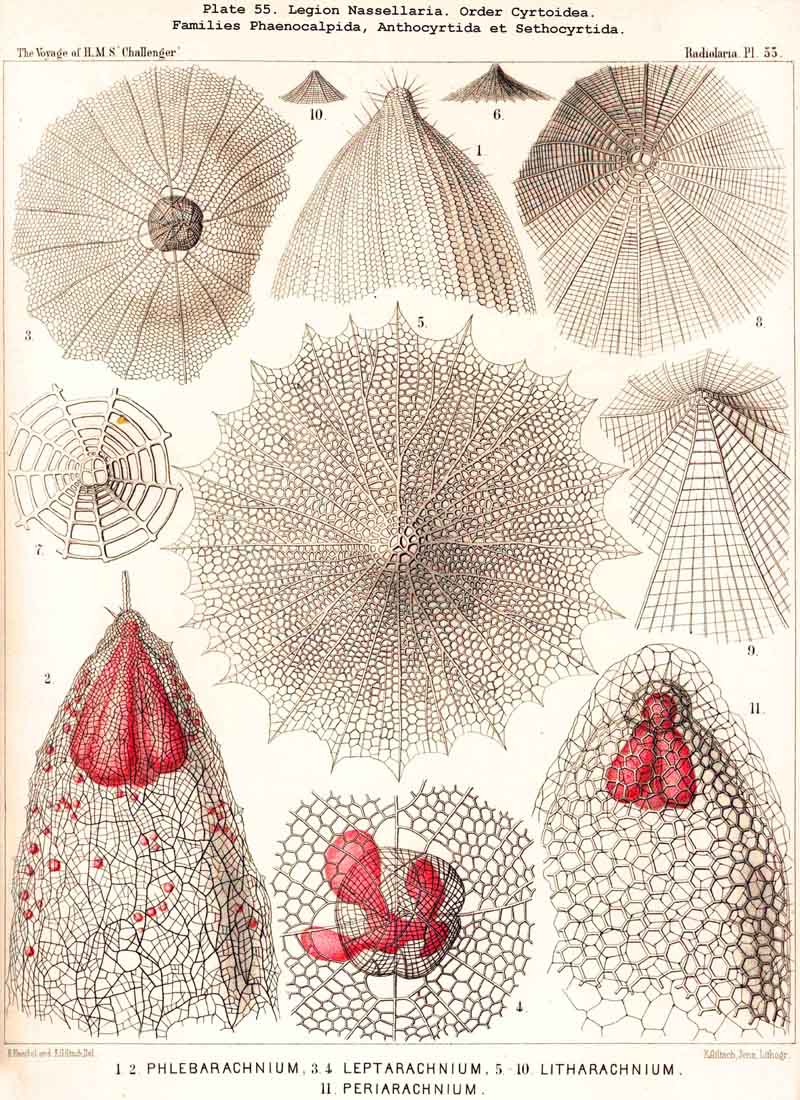{kind=link}
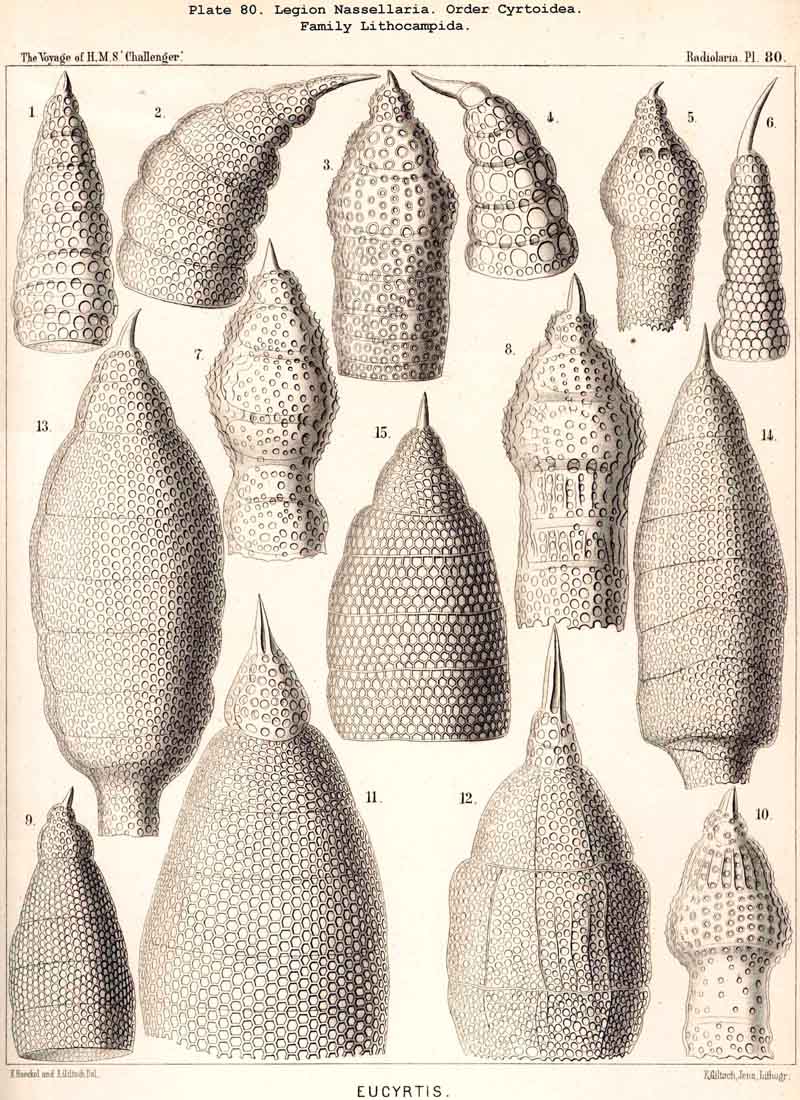{kind=link}
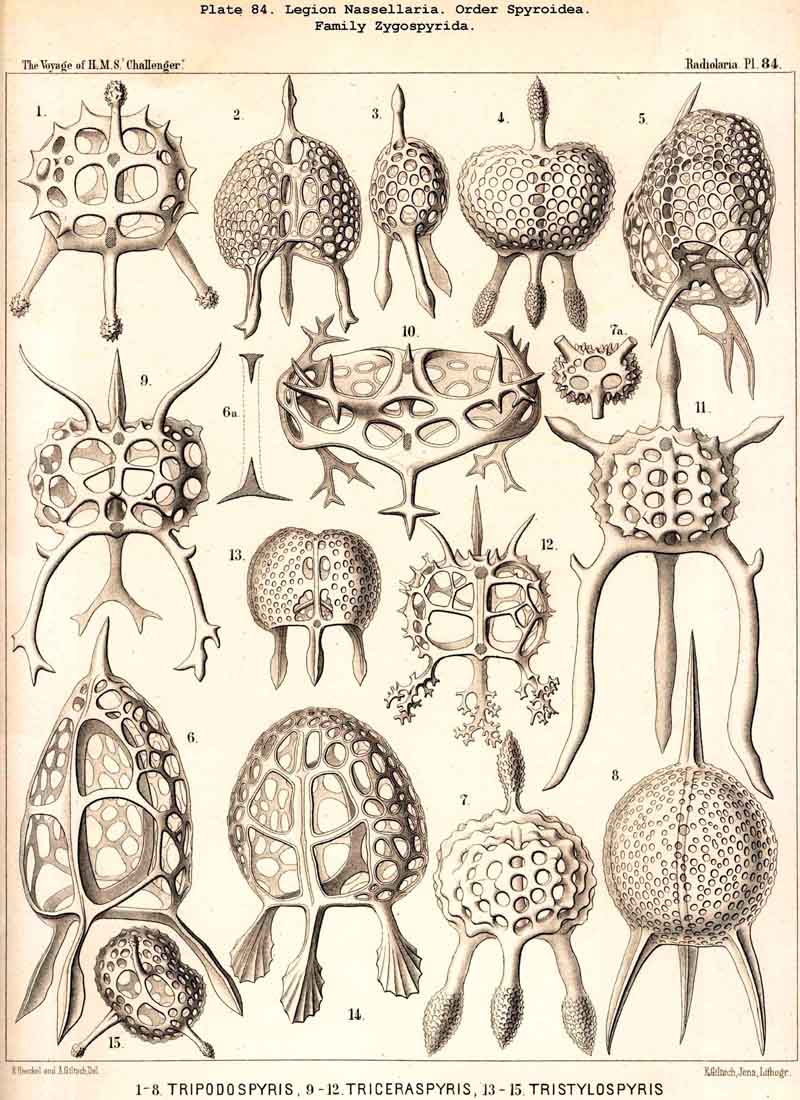{kind=link}
{kind=link}
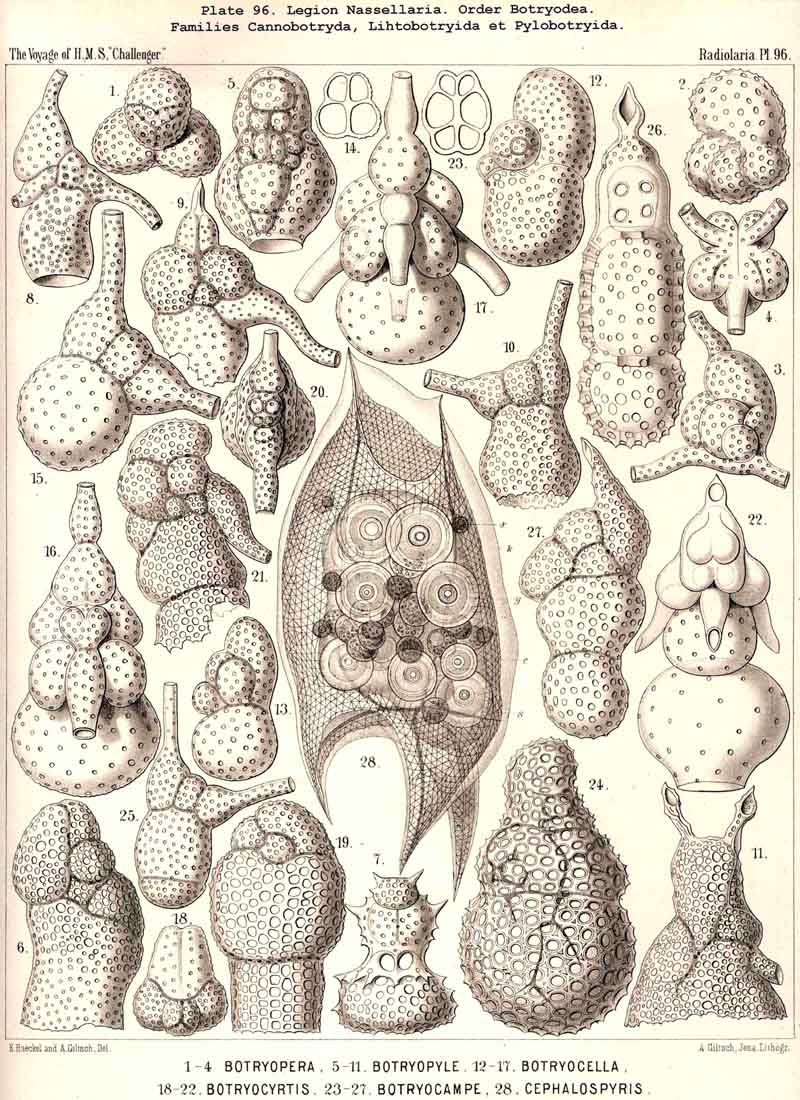{kind=link}
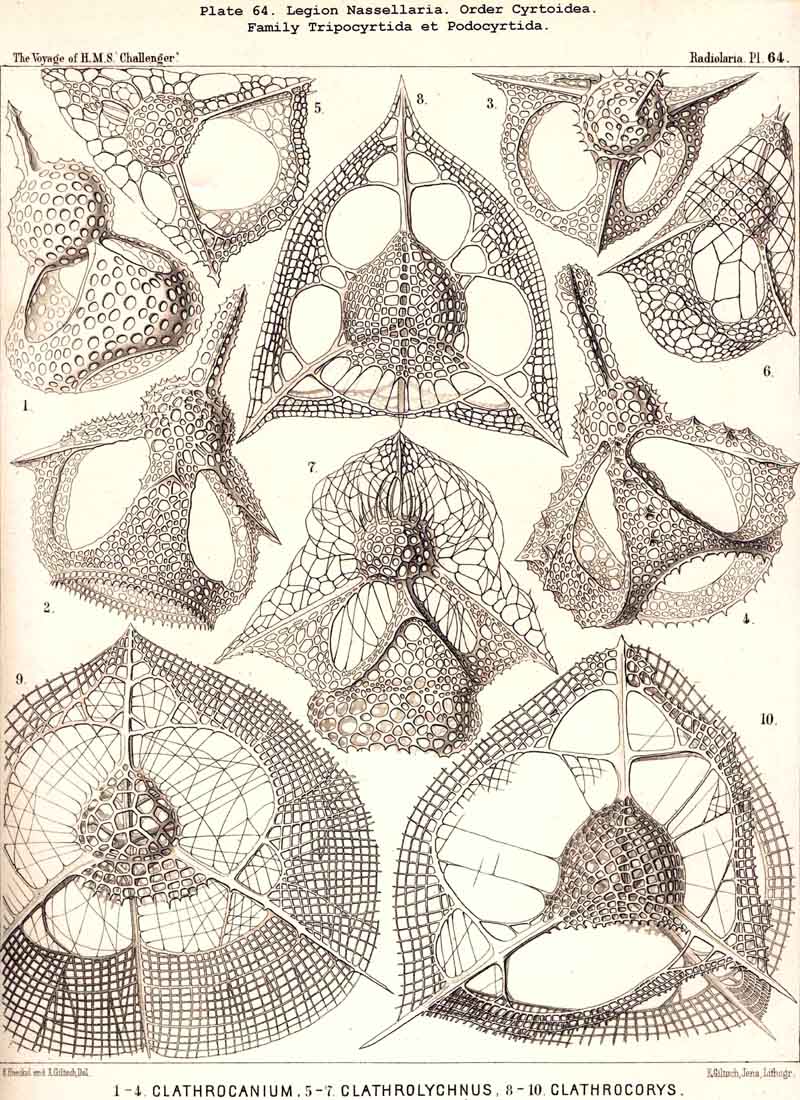{kind=link}
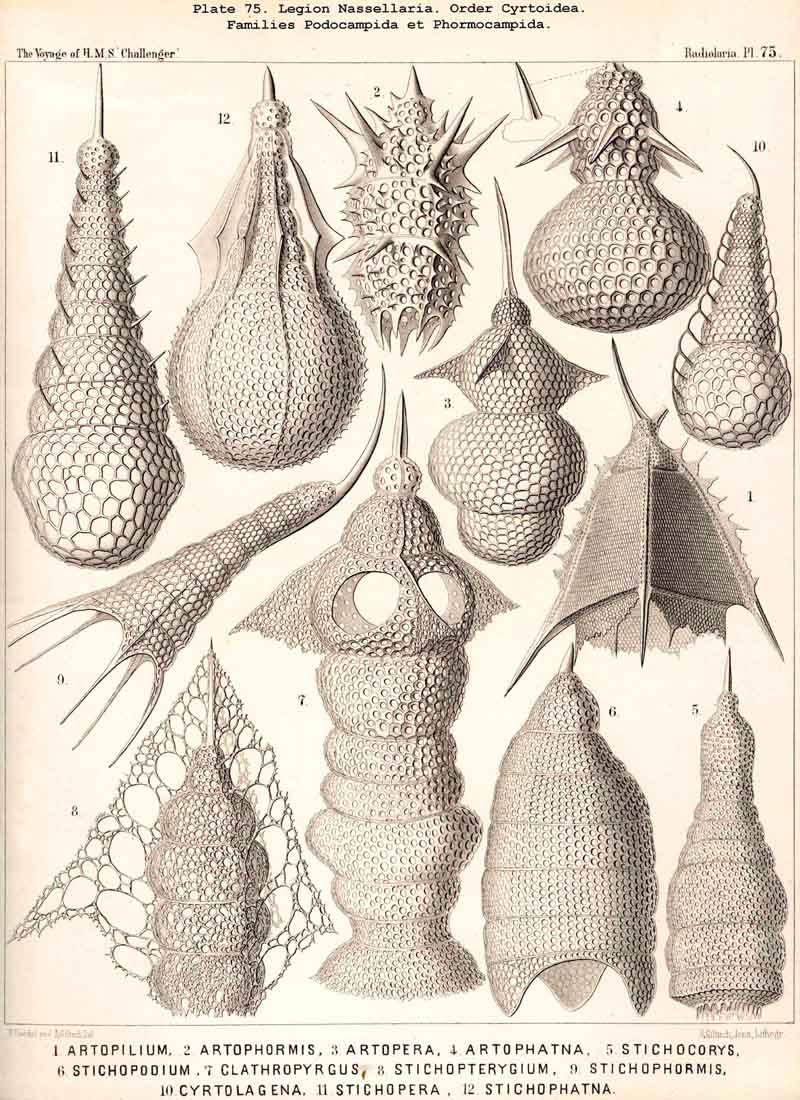{kind=link}
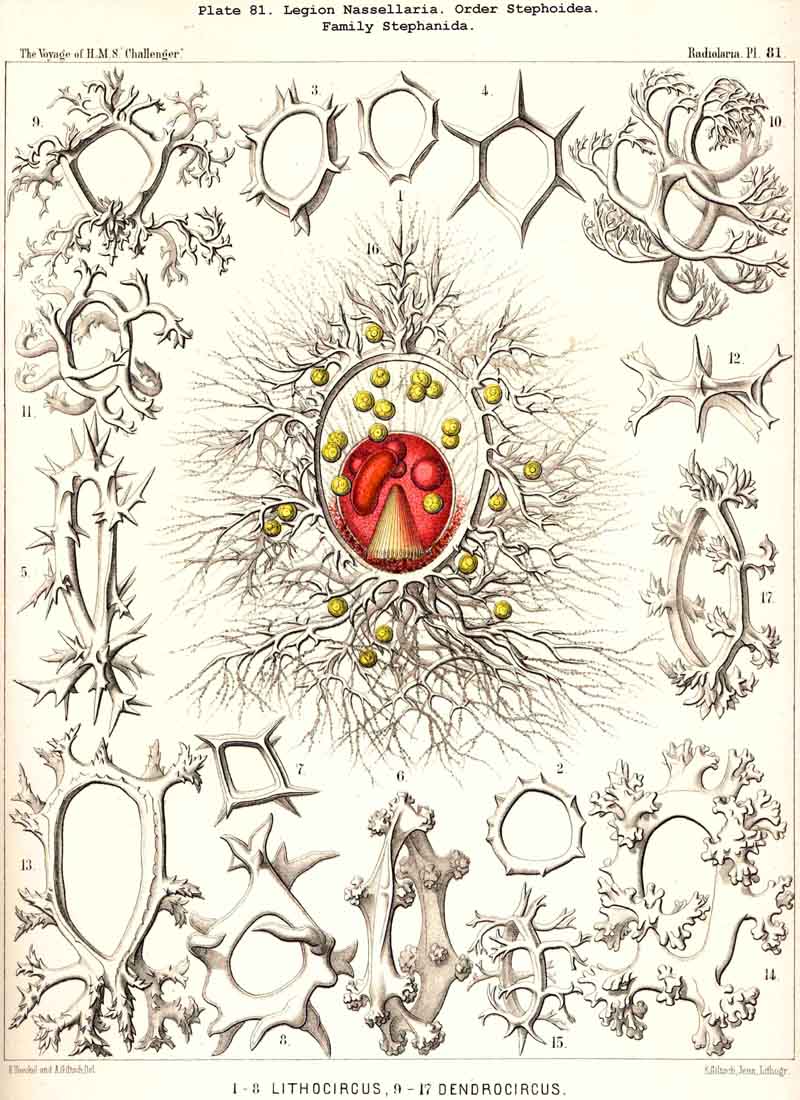{kind=link}
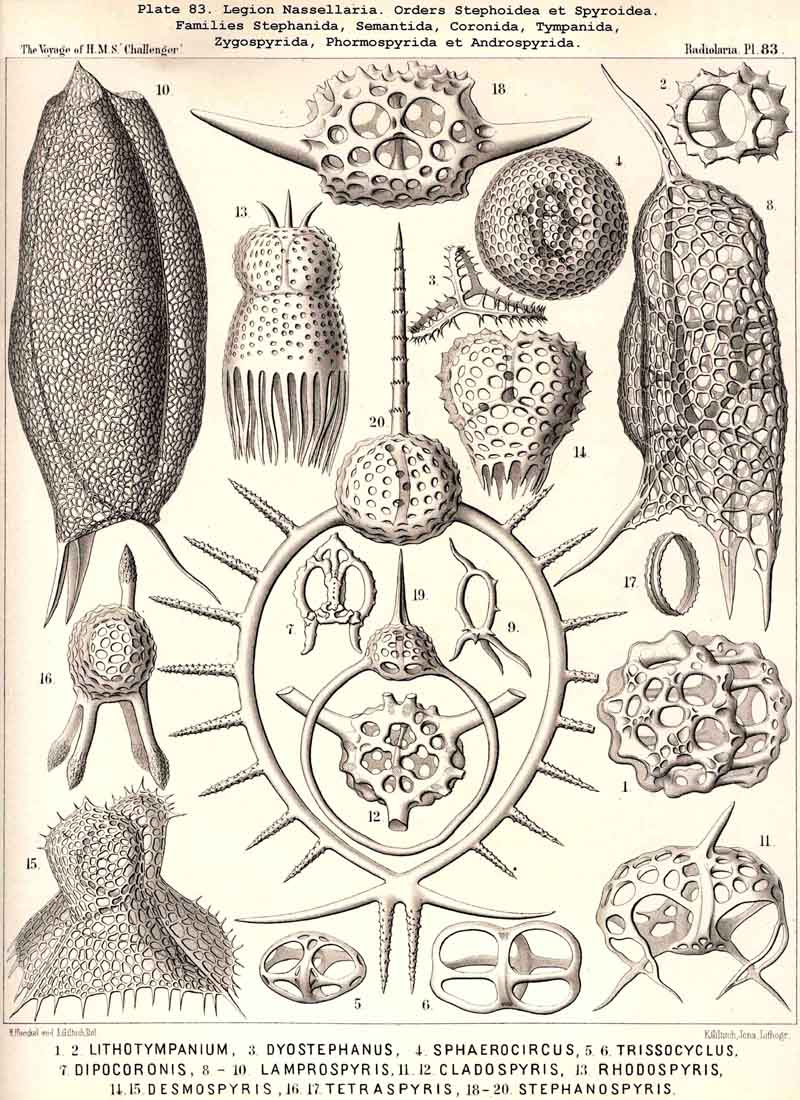{kind=link}
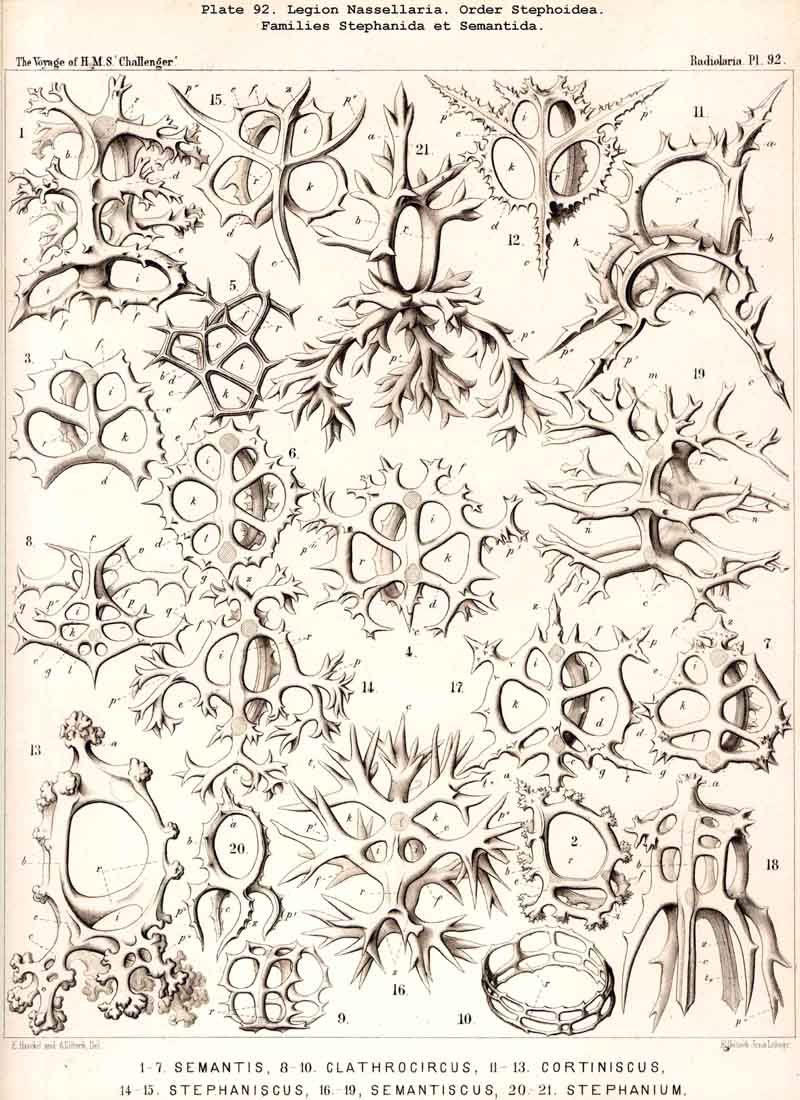{kind=link}
{kind=link}
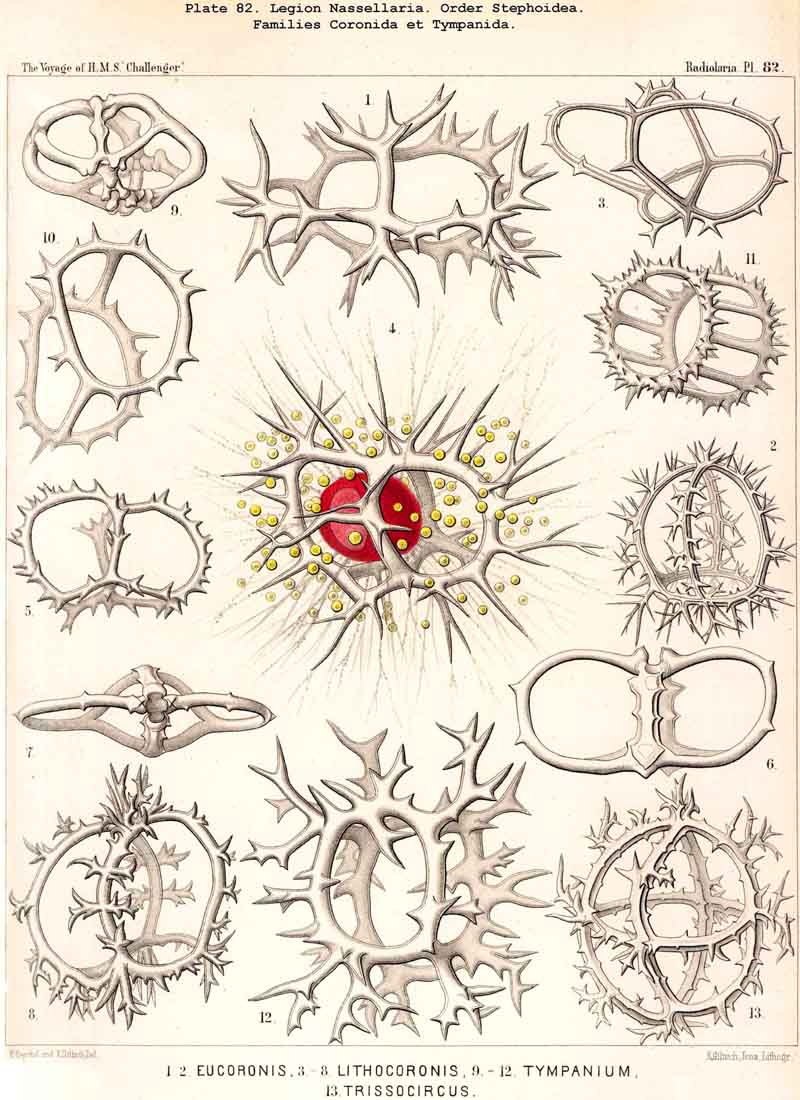{kind=link}
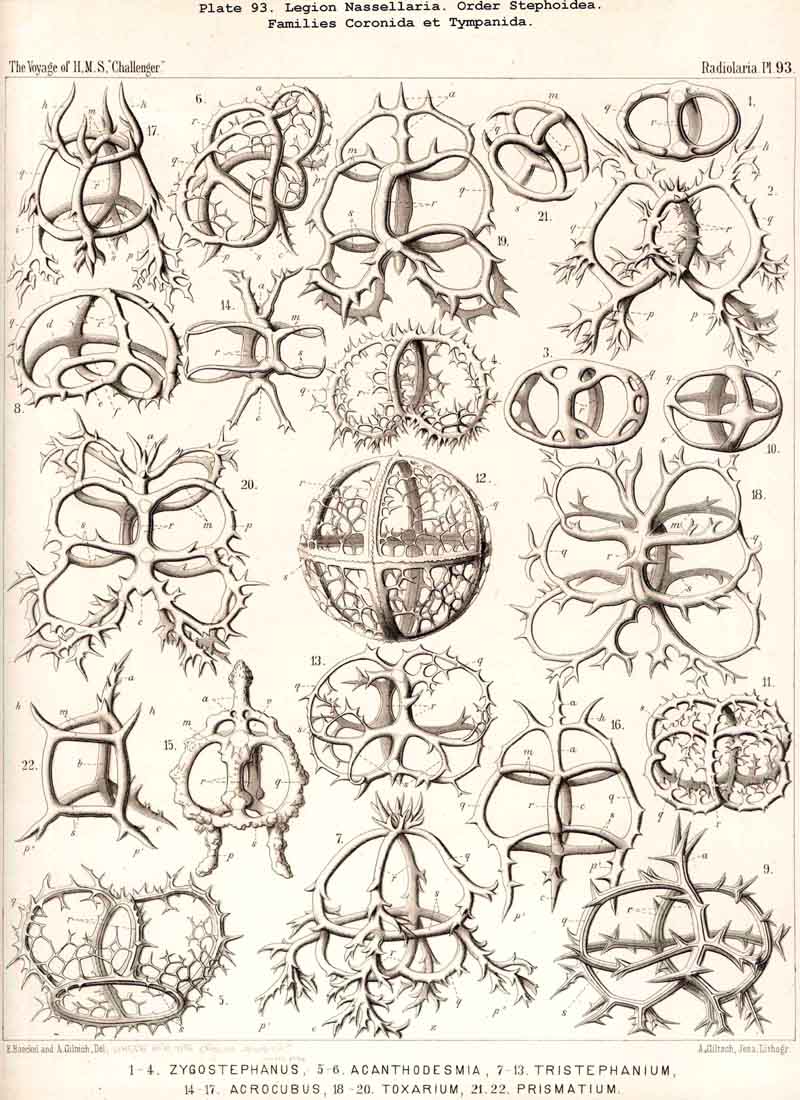{kind=link}
{kind=link}
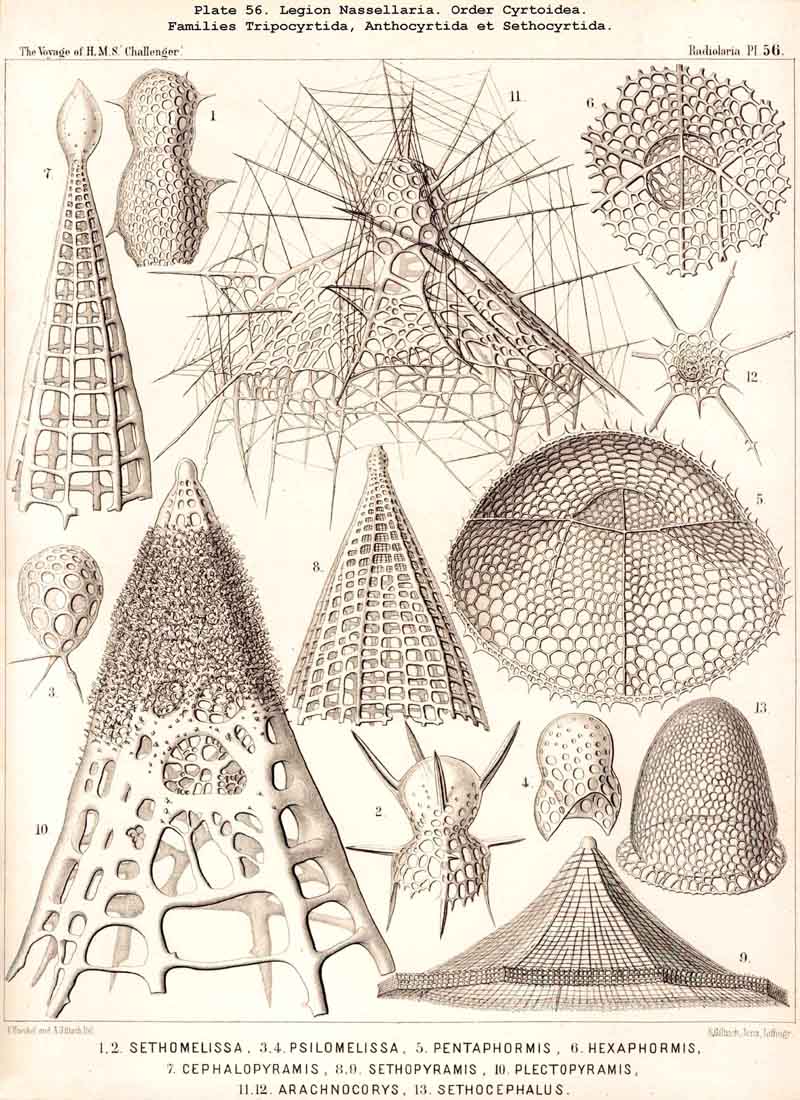{kind=link}
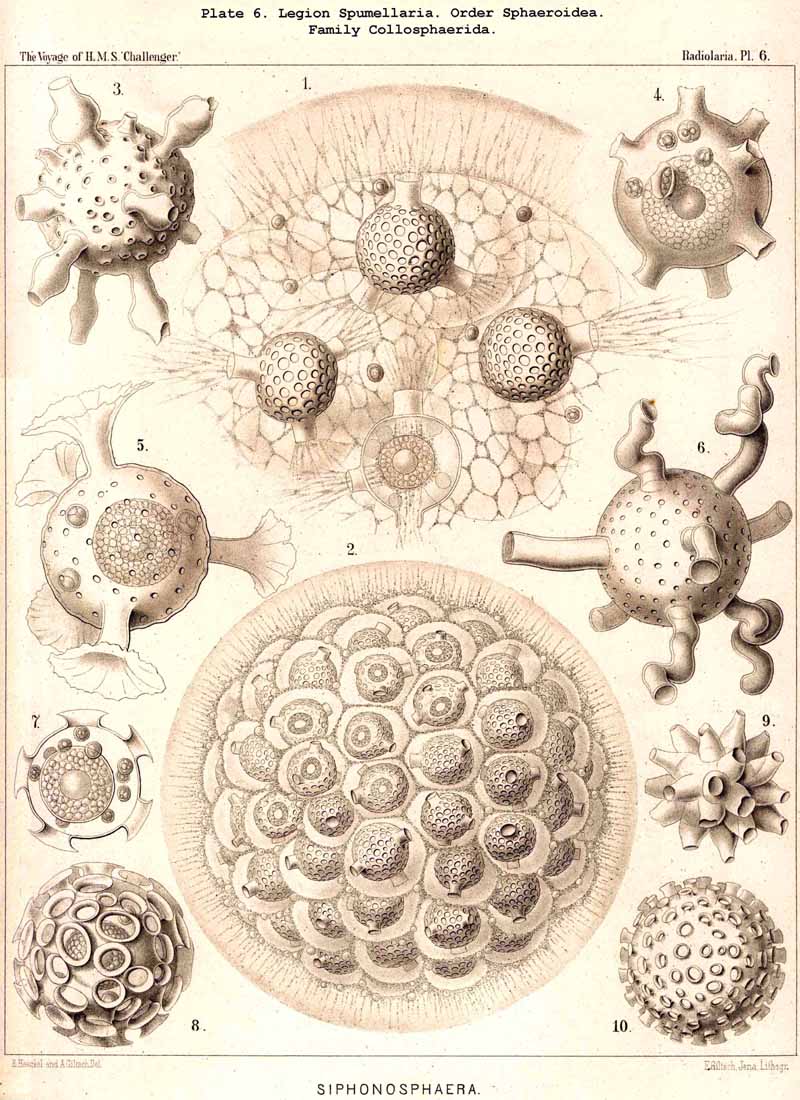{kind=link}
{kind=link}
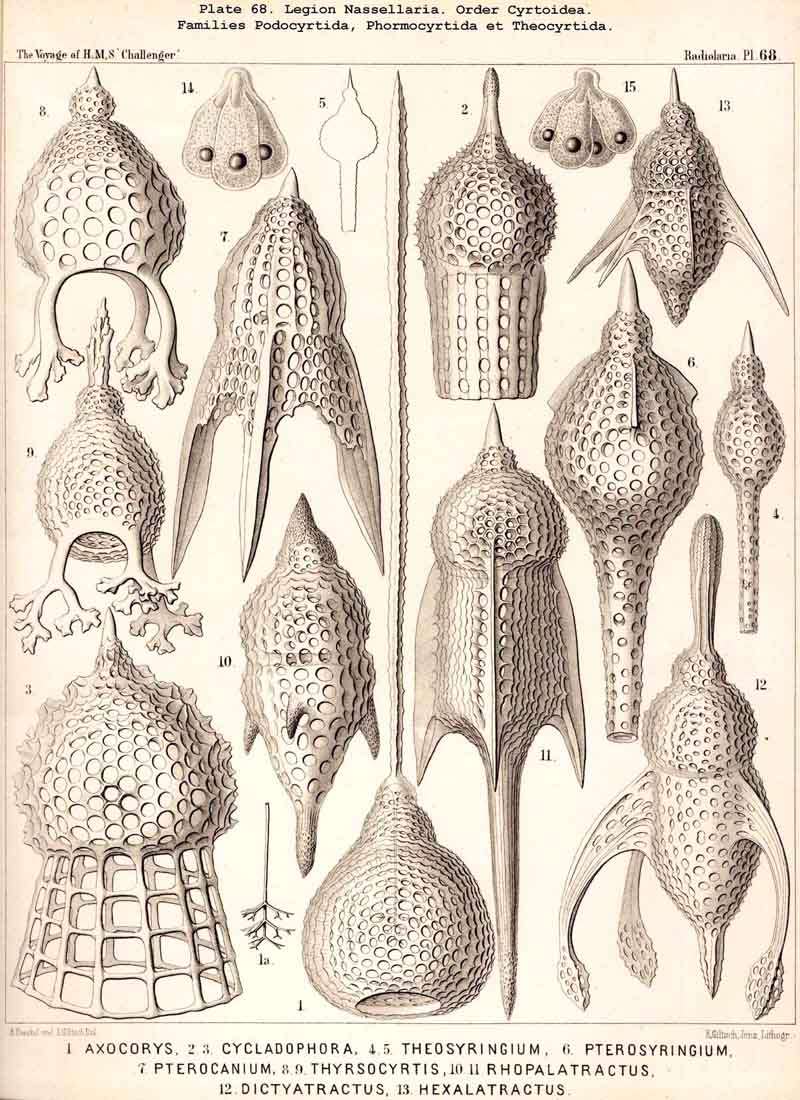{kind=link}
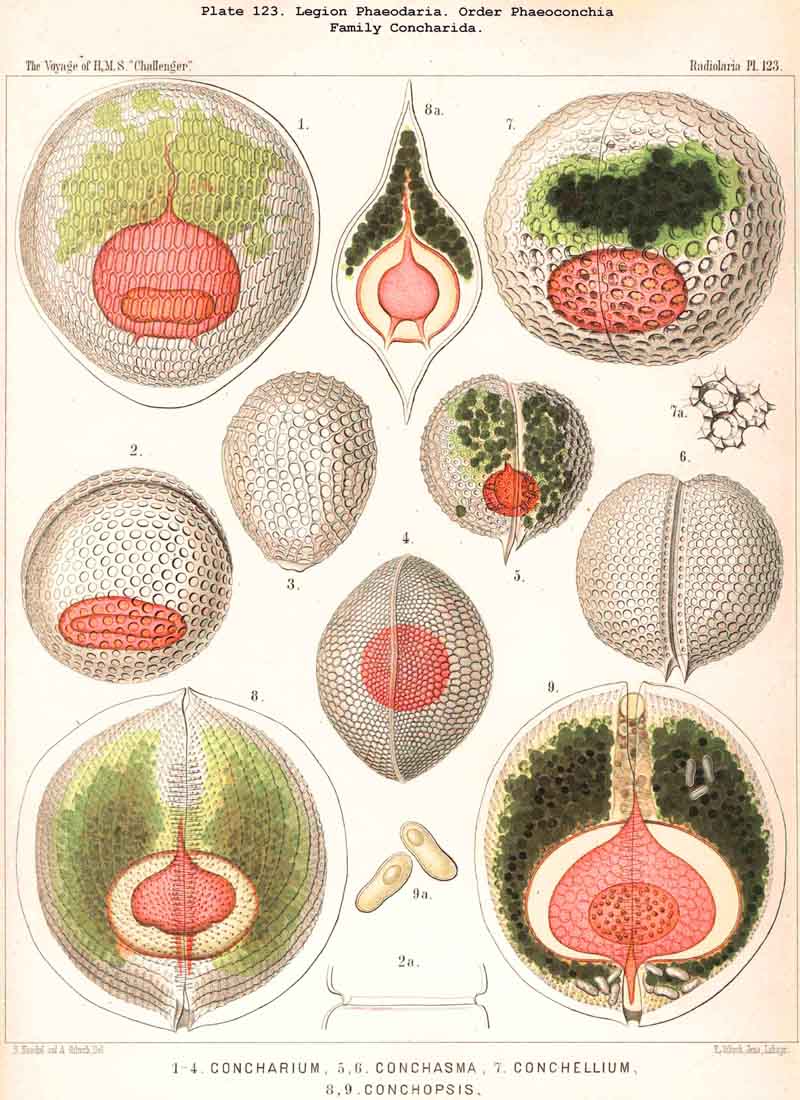{kind=link}
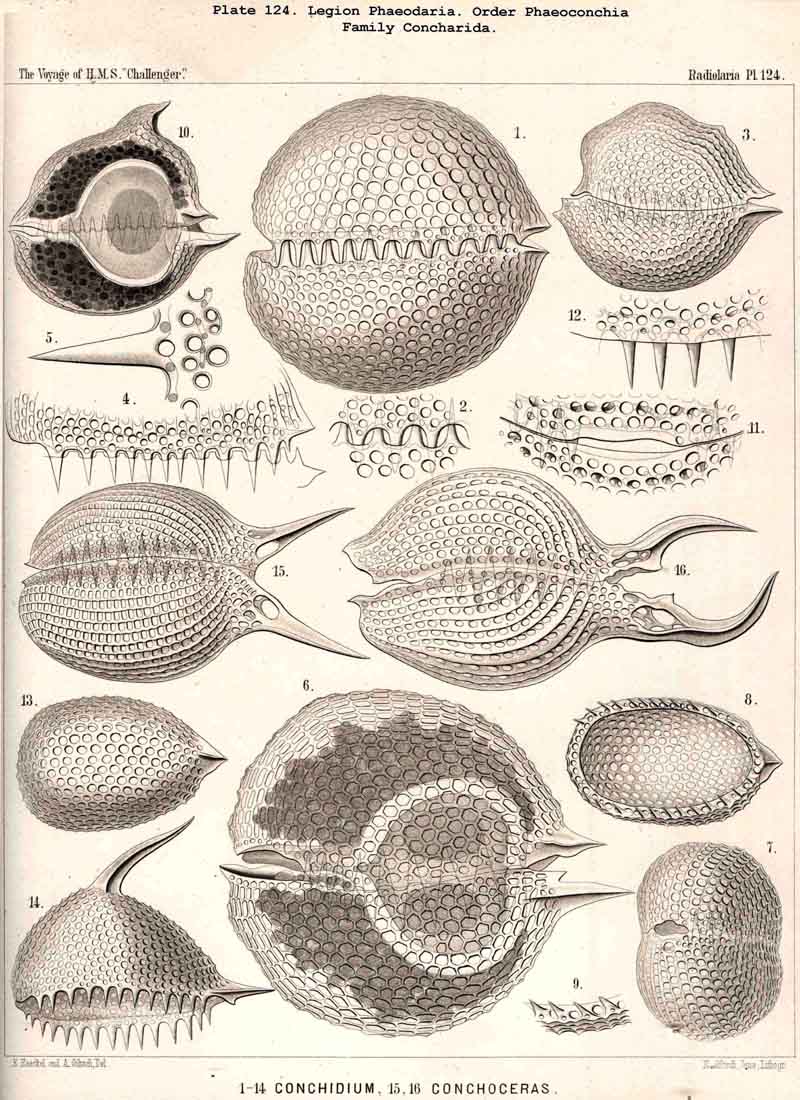{kind=link}
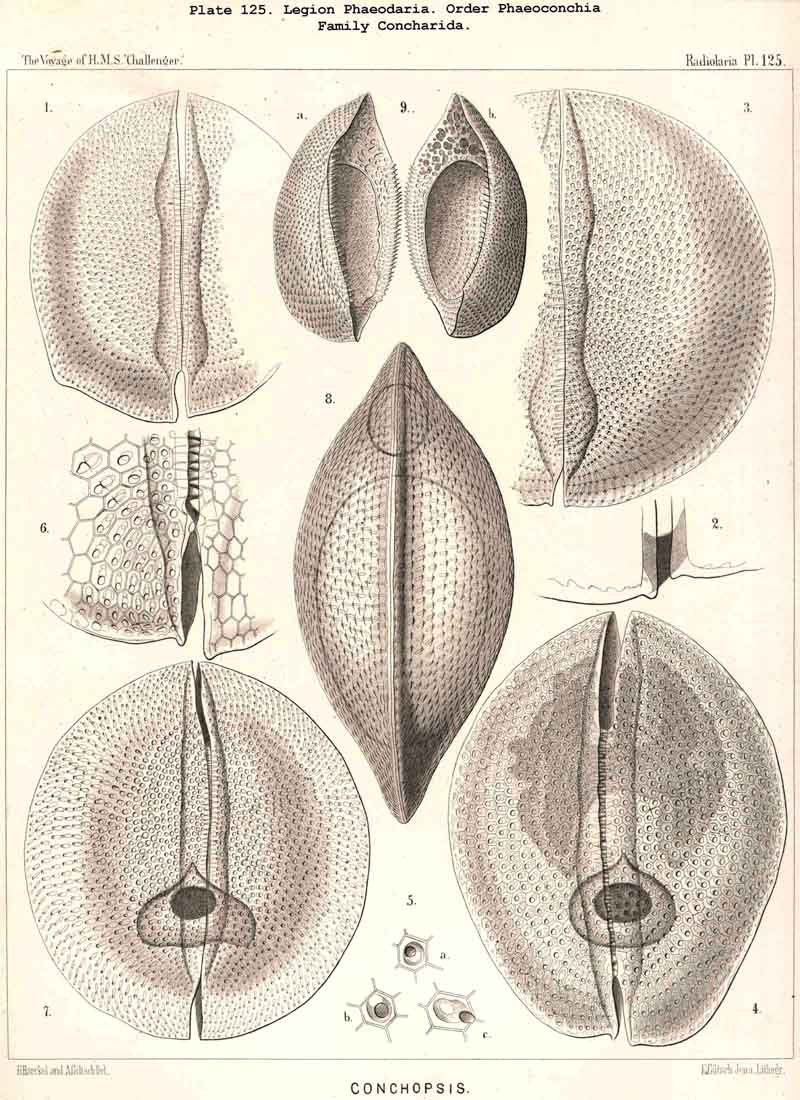{kind=link}
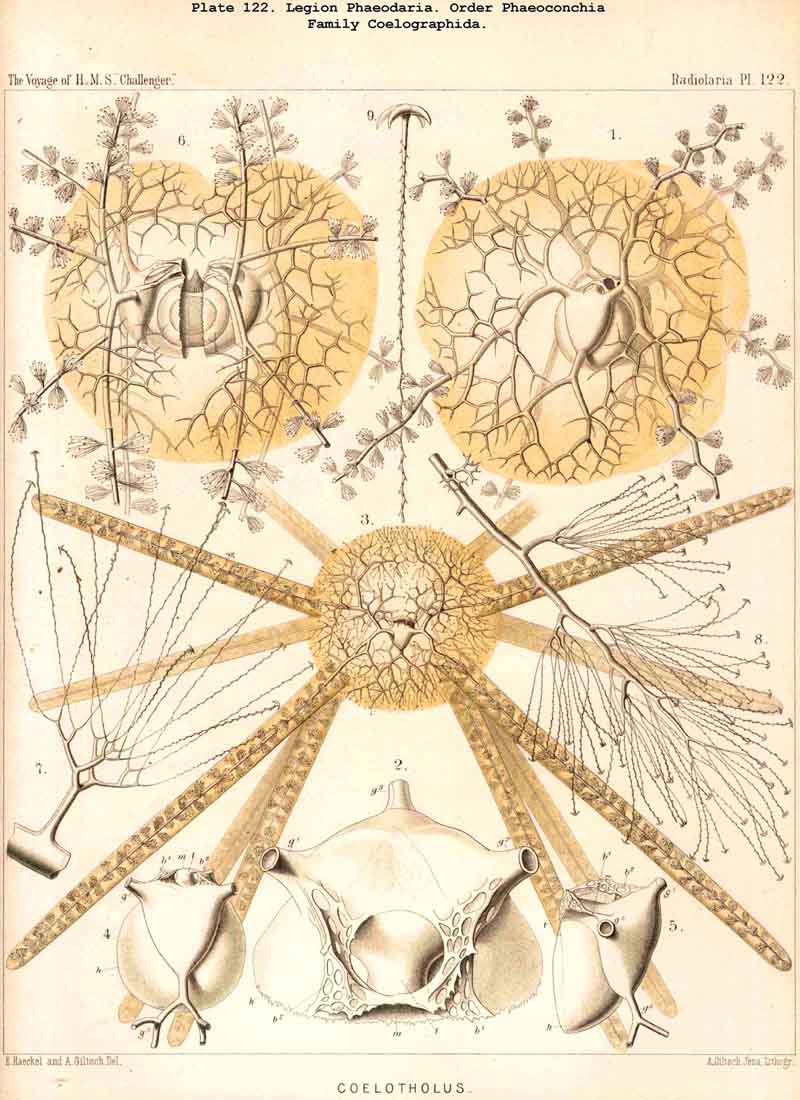{kind=link}
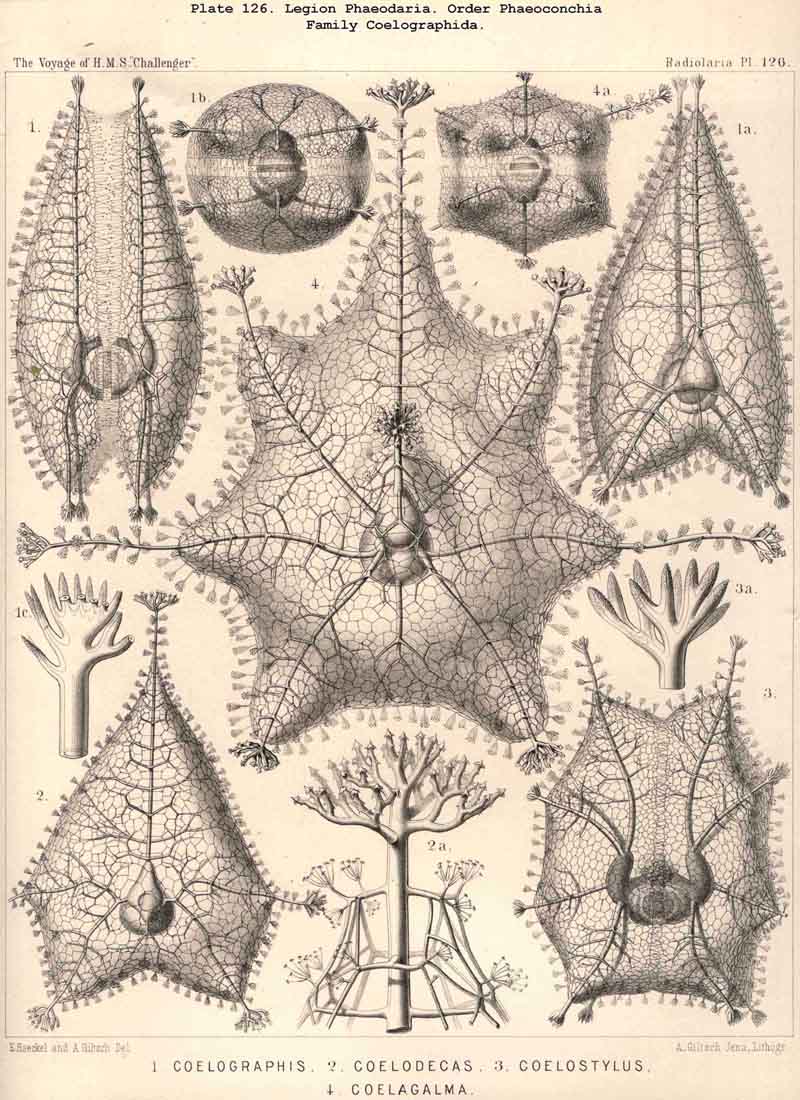{kind=link}
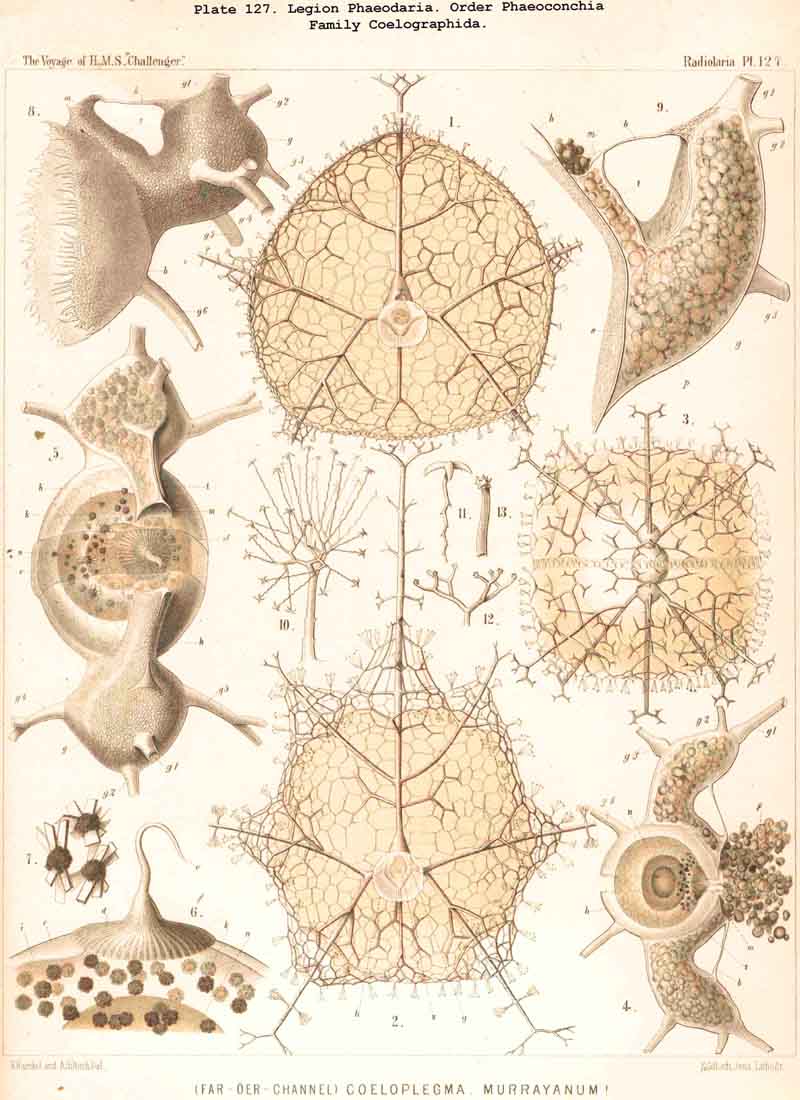{kind=link}
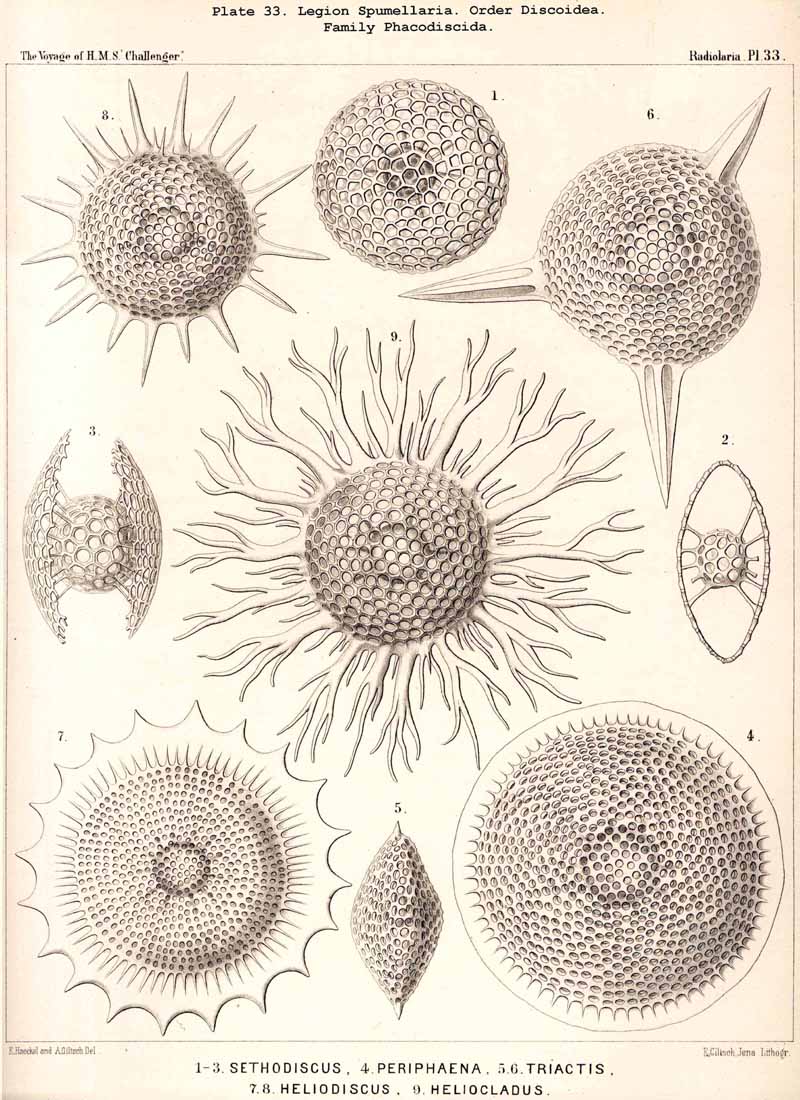{kind=link}
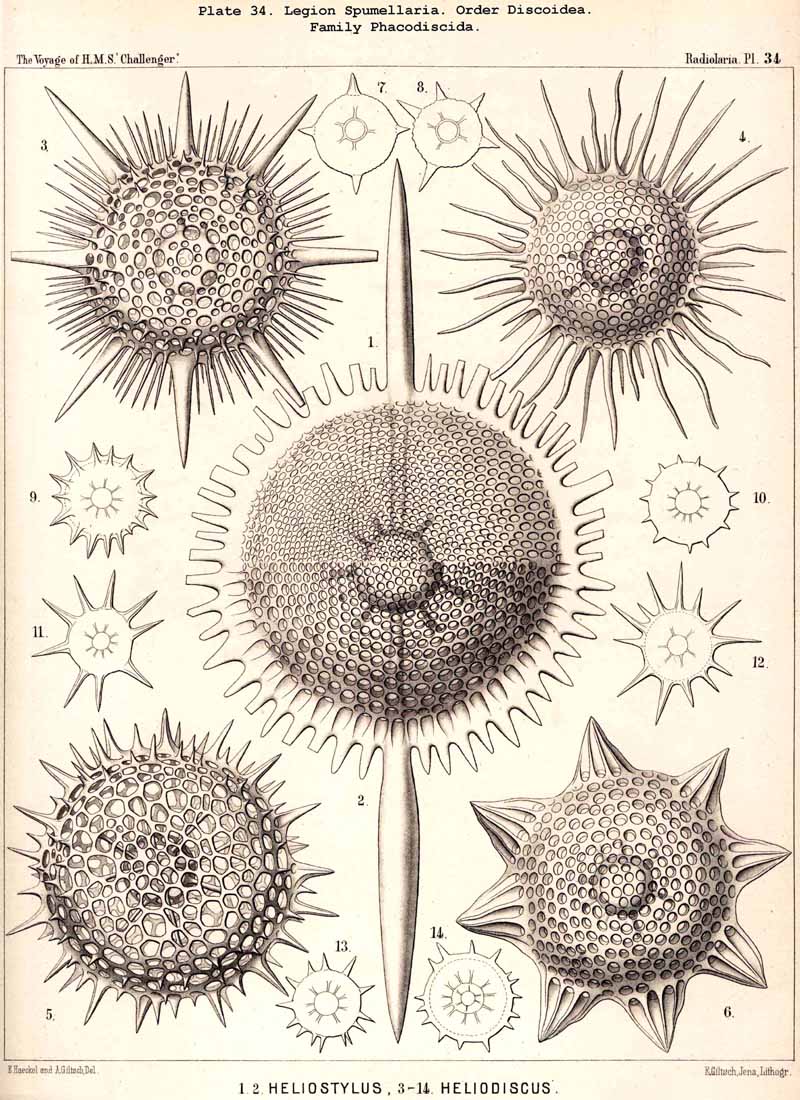{kind=link}
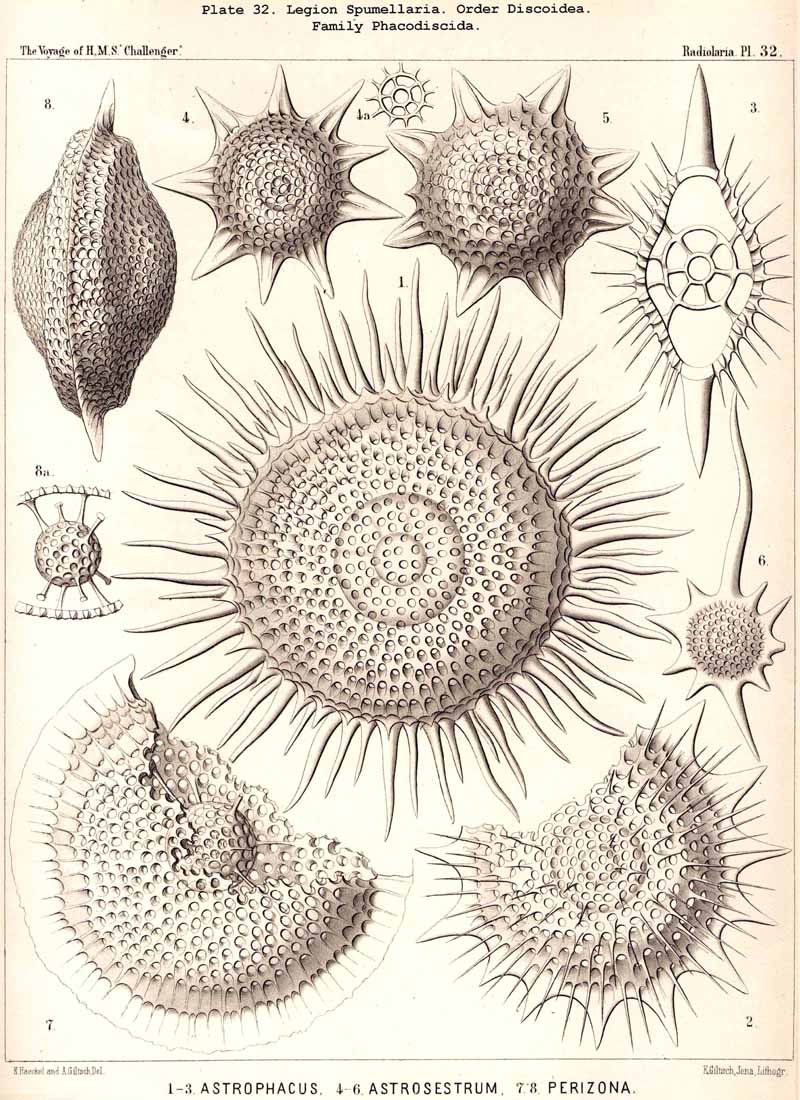{kind=link}
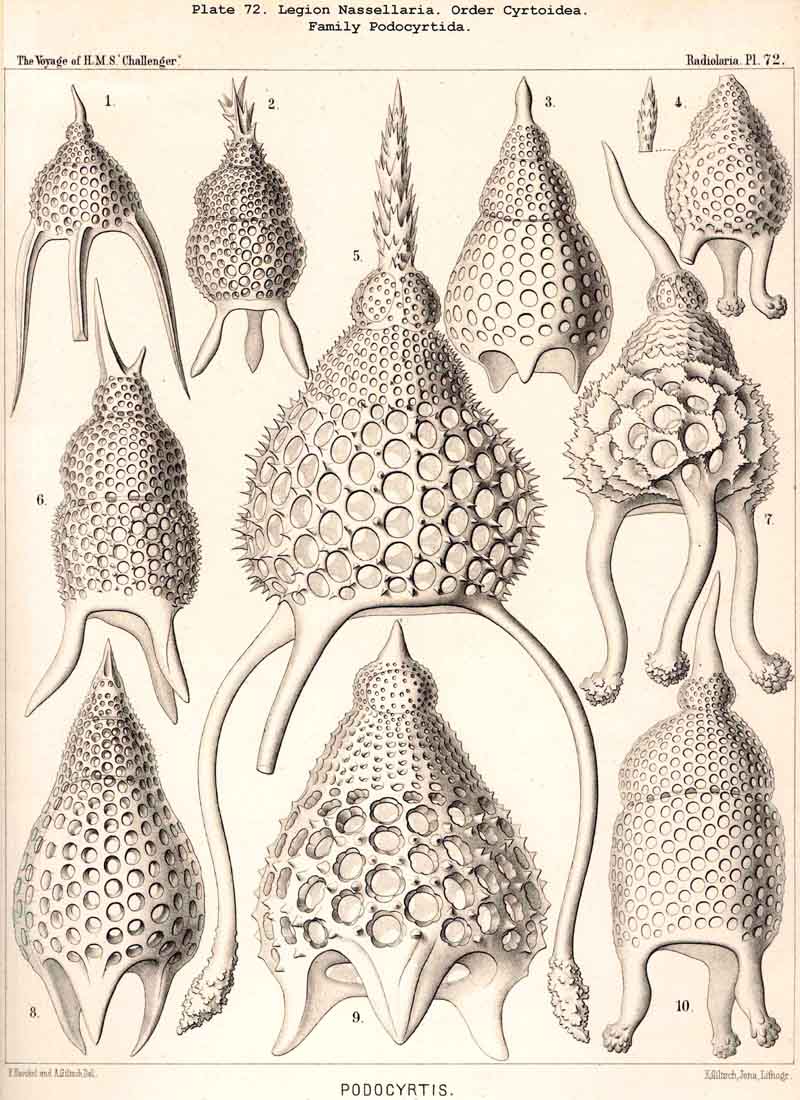{kind=link}
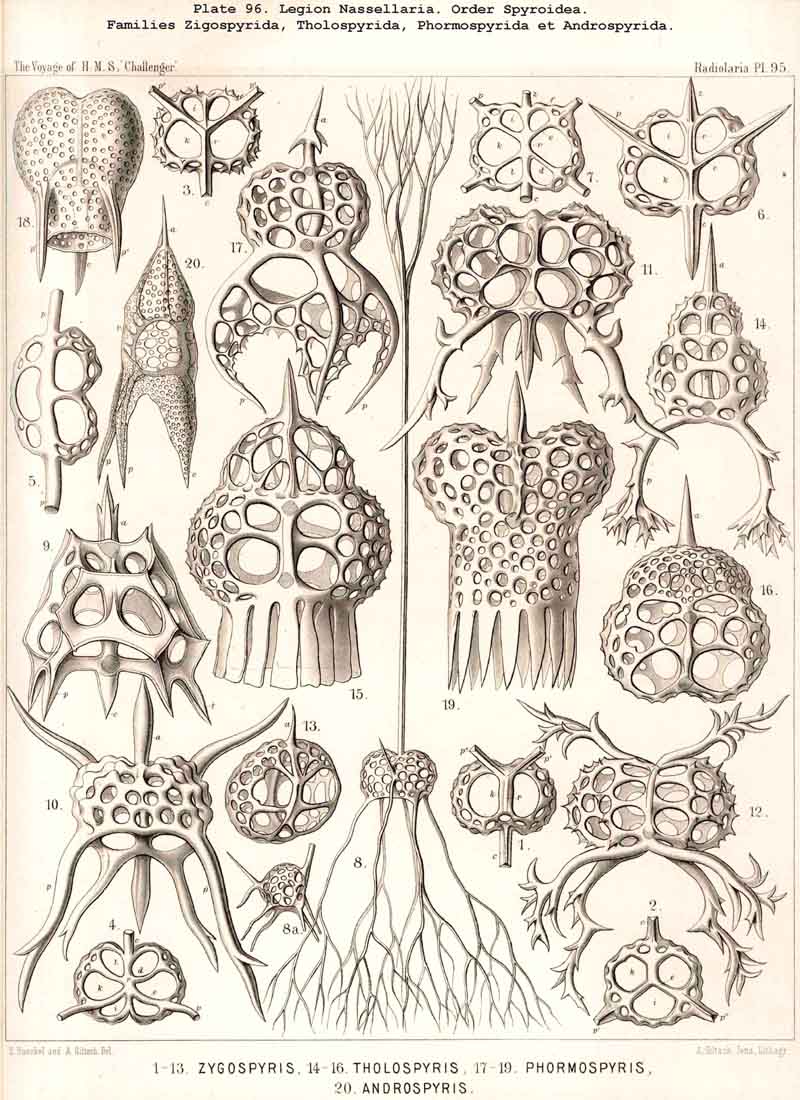{kind=link}
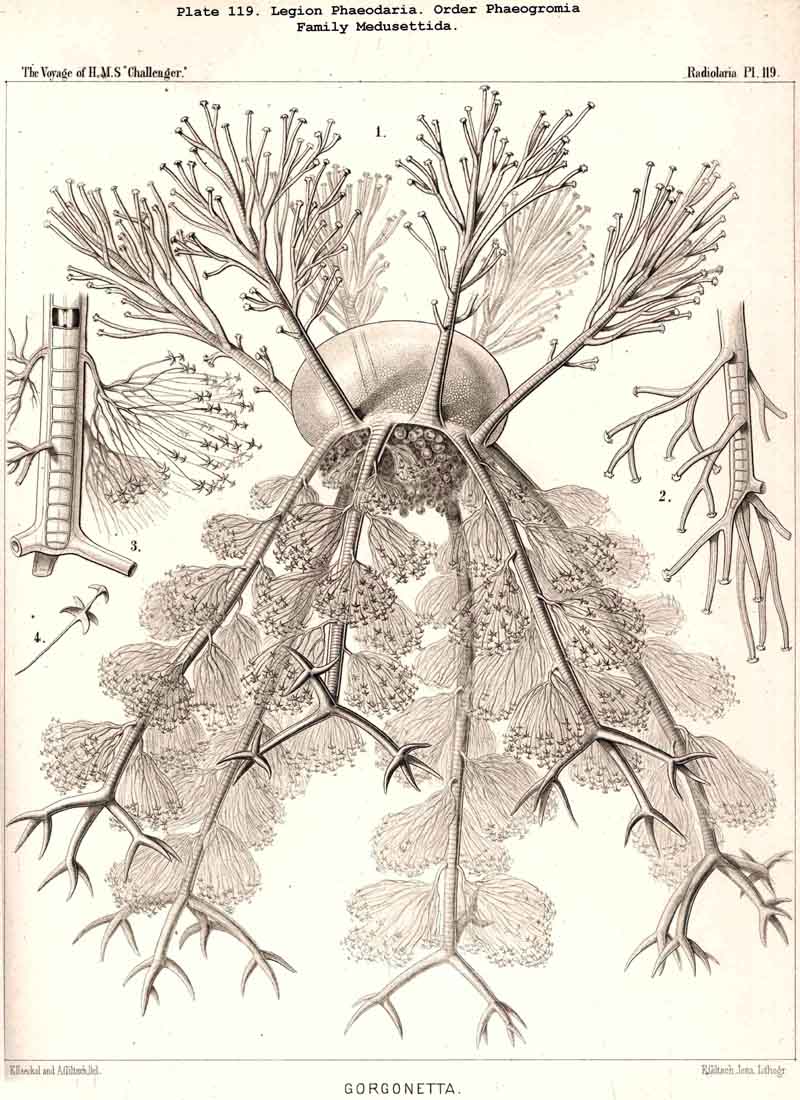{kind=link}