A SKETCH OF OUR KNOWLEDGE OF THE DEVELOPMENT OF THE RADIOLARIA
IN THE YEAR 1884.
CHAPTER V.ONTOGENY OR INDIVIDUAL DEVELOPMENT.
(§§ 141-152.)
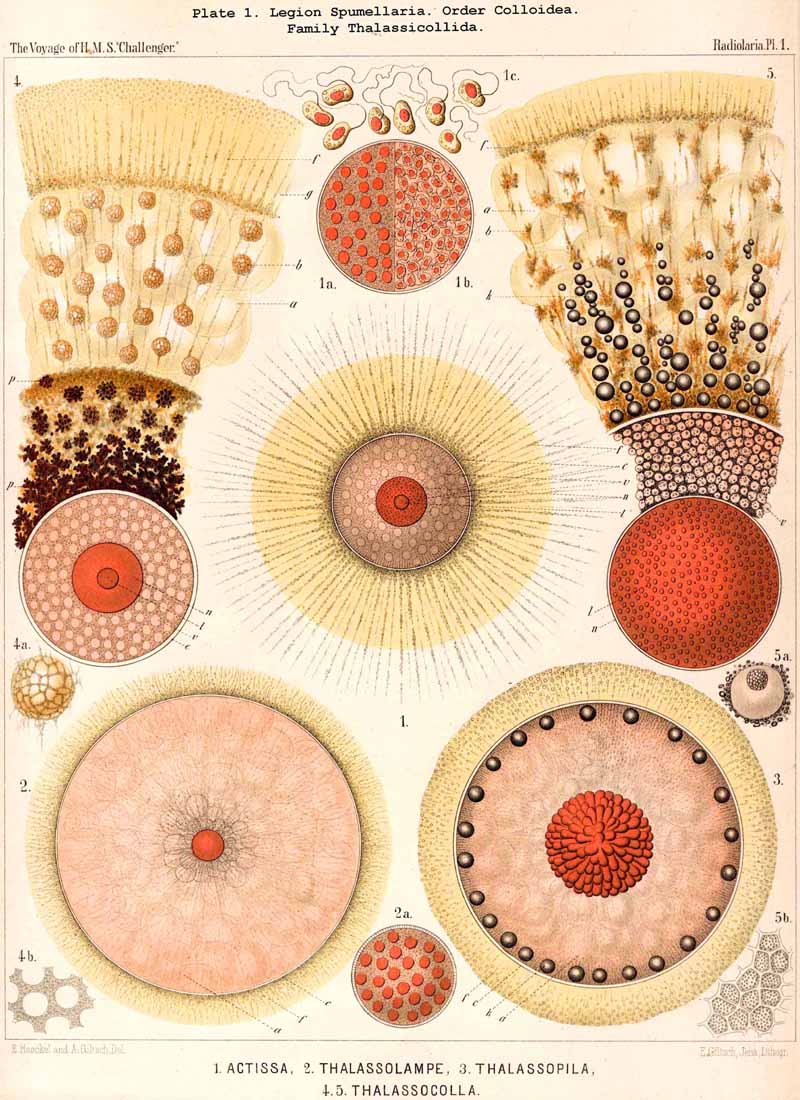
141. Individual Developmental Stages.-The germinal history of the Radiolaria presents great obstacles to direct observation, and hence is very incompletely known. The fragmentary observations, however (having been made on Radiolaria of very various groups and supplemented by comparative anatomical considerations), allow us to draw a general picture of the essential developmental processes in this great class. It may probably be assumed that in all Radiolaria, after maturation, the central capsule discharges the function of a sporangium, and its contents are broken up into numerous flagellate swarm-spores (zoospores). After these flagellate swarm-spores (resembling Astasia) have emerged from the ruptured central capsule, they probably pass over into a Heliozoan-stage (Actinophrys) and then after the formation of a jelly-veil into the condition of Sphaerastrum. Afterwards, when a membrane is formed between the outer jelly-veil and the inner nucleated cell-body, an Actissa-stage arises, which exhibits in its simplest form the differentiation of the spherical unicellular body into the central capsule and calymma. Actissa thus represents both ontogenetically and phylogenetically the primitive condition of the Radiolarian organism, and may thus be regarded as the point of departure of all other forms.
142. The Astasia-Stage.-The formation of flagellate zoospores in the mature central capsule is probably to be regarded as the common form of individual development in all Radiolaria ; since the whole contents are utilised in the formation of these swarm-spores, and since the extracapsulum takes no share in the process and perishes after they are evacuated, the central capsule may be regarded as a sporangium (see note A, below). The zoospores of the Radiolaria generally arise in the following way:- the nucleus of the unicellular organism, sometimes early, sometimes late (and in several different ways, §§ 63-70) breaks up into numerous small nuclei, and each of these surrounds itself with a small portion of the endoplasm. Very often, perhaps generally, this endoplasm contains one or several fat-granules and sometimes also a small oblong Crystal ; from the protoplasm of the small roundish or ovoid cells protrudes one or more vibratile flagella. The fully developed spores, which commence their vibrations even within the central capsule, emerge when it ruptures, and swim about freely in the surrounding water by means of the flagellum. At this stage of its existence the young Radiolarian represents essentially the simplest form of the Flagellata, such as Astasia or Euglena ; the unicellular body is for the most part ovoid or subcyhindricah, sometimes fusiform or reniform, usually from 0.004 to 0.008 mm. in diameter (Pl. 1, fig. 1c ; Pl. 129, fig. 11). In the anterior part of the flagellate cell, immediately behind the base of the flagellum, lies a homogeneous, spherical nucleus, whilst in the posterior part are usually several small fat-granules and often also a small oblong crystal (hence the name "crystal-spore," "Krystall Schwärmer"). The number of vibrating flagella, which are extremely long and fine, seems to be variable, usually one, sometimes two, occasionally perhaps three, or even four or more (see note B).
A. The formation of the motile spores in the central capsule was first observed by J. Müller in Acanthometra(1856, L. N. 10, p. 502), then by A. Schneider in Thalassicolla (1858, L. N. 13, p. 41), and finally by myself in Sphaerozoum (1859, L. N. 16, p. 141). These older observations were, however, incomplete, for the origin of the motile corpuscles from the contents of the central capsule was not observed. The first complete and detailed observations upon the formation of spores in the Radiolaria were published in 1871 by Cienkowski (L. N. 22, p. 372, Taf. xxix.); they relate to two different Polycyttaria, Collosphaera and Collozoium. These investigations were supplemented by R. Hertwig on Collozoiumand Thalassicolla (1876, L. N. 26, pp. 28, 43, &c.); on Collozoum. he made the important discovery that the Polycyttaria form two kinds of spores, one with and the other without crystals, and that the latter are divided into macrospores and microspores (compare the chapter on "Reproduction," §§ 212-216). Quite recently Karl Brandt has confirmed these observations, and has extended them to all the genera of Polycyttaria (1881, L. N. 38, p. 393, and 1885, loc. cit.).
B. The number of flagella, projecting from each spore, is very difficult to determine, owing to their extraordinary length and slenderness. It appeared to me that in the majority of those Radiolaria whose spores I investigated only a single flagellum could be demonstrated with certainty, although sometimes two, springing from a common base, seemed to be present. Compare the chapter on "Reproduction," (§ 215) and the recent work of Karl Brandt on Sphaeozoea (1885, L. N. 52, pp. 145-174).
143. The Actinophrys-Stage.-The fate of the flagellate zoospores which emerge from the mature central capsule of the Radiolaria has not hitherto been decided by actual observation ; all attempts to rear the swarming zoospores have been in vain, for they have soon died. From what we know, however, of the comparative morphology of the Protista, the hypothesis is fully justified, that between the Astasia-stage of the flagellate swarm-spores, and the well-known Actissa-stage of the simplest Radiolaria, there lies an intermediate developmental stage, which may be regarded as being essentially the simplest Heliozoan form, Actinophrys or Heterophrys. The swarm-spore is very probably converted directly into a simple floating Heliozoon by its elongated or ovoid body becoming spherical and by fine pseudopodia protruding all round instead of a single flagellum ; the nucleus at the same time assuming a central position.
144. The Sphaerastrum-Stage.-The Actinophrys-stage of the young Radiolaria, which proceeds immediately from the flagellate zoospore, is probably connected with the Actissa-stage by an intermediate form, which may be regarded as a simple skeleton-less Heliozoon with a jelly-veil ; a well-known example of such a form is Sphaerastrum (in the solitary, not the social condition) and Heterophrys. This important intermediate form has arisen from the simple Actinophrys-stage by the excretion of an external structureless jelly-veil, such as is formed in many other Protista (e.g., in the encystation of many Infusoria). The young Radiolarian in this second Heliozoon-stage becomes a simple cell with pseudopodia radiating on all sides ; its body consists of three concentric spheres, the central nucleus, the protoplasmic body proper, and the surrounding calymma or jelly-veil. When a firm membrane is developed between the last two spheres this Sphaerastrum-stage passes over into the Actissa.
The gap in our empirical knowledge which still exists between the flagellate
stage (§ 142) and the simplest Radiolarian stage (Actissa, §
145), can be filled hypothetically only by the assumption of several Heliozoon-stages
following one upon another. It is possible also that the capsule-membrane
is not formed between the endoplasm and exoplasm (as here supposed), but
that the membrane was formed first outside the cell and the extracapsulum
subsequently secreted around it.
145. The Actissa-Stage.-The first SPUMELLARIAN genus, Actissa, is not only the simplest form actually observed among the Radiolaria, and the true prototype of the whole class, but also the simplest form under which the Radiolarian organisation can be conceived. It is therefore extremely probable that Actissa not only forms the common stem-form of the whole class in a phylogenetic sense, but is also its common ontogenetic or germinal form. Probably in all Radiolaria the Sphaerastrum-stage develops immediately into the typical Actissa-stage, by the formation of a firm membrane between the protoplasmic body of the spherical Heliozoan cell and its jelly-veil. Thus arises the characteristic central capsule, which is wanting in the nearly related Hehiozoa. It is further probable that all Radiolaria in their early stage will so far conform to the state of things in Actissa as to have the capsule-membrane of the spherical skeletonless cell perforated everywhere by fine pores. This structure is retained in all SPUMELLARIA, whilst in the other three legions those structural relations of the capsule which are characteristic of each develop from the Actissa-stage.
146. The Ontogeny of the Spumellaria.-In the simplest case the individual development in the SPUMELLARIA ceases with the Actissa-stage. In all other genera of this legion diverging forms proceed from this; of which the different growth of the three dimensive axes on the one hand (§§ 44, 45), and the differentiation of the various parts of the unicellular organism with the formation of the skeleton on the other, are of pre-eminent significance. Even in the varying growth of the central capsule in the different dimensions of space in the skeletonless Colloidea, four different modes may be distinguished, which further, in the corresponding development of the skeleton, furnish the basis for the origin of the four orders of Sphaerellaria. The most primitive and simplest form of growth, equal extension in all directions, is found in the spherical central capsule and the concentric spherical skeletons (Procyttarium, Sphaeroidea). When the growth of the central capsule proceeds more rapidly in the direction of the vertical main axis than in any other direction, the ellipsoidal or cylindrical central capsule (Actiprunum) arises, and the vertically elongated skeleton of the Prunoidea, which is derived from it. When, on the contrary, the growth of the central capsule and lattice-shell is less in the direction of the vertical main axis than in any other direction, the lenticubar or discoid central capsule (Actidiscus) arises, and the corresponding lenticular shell of the Discoidea. Finally, even quite early in many SPUMELLARIA, the growth of the central capsule and of the corresponding lattice-shell in the three dimensive axes is different, and hence arise the lentelliptical forms whose geometrical type is the triaxial ellipsoid or the rhombic octahedron (Actilarcus, Larcoidea). Thus the origin of the four orders of Sphaerellaria is simply explained by a varying growth in the different dimensive axes. The primary (innermost) lattice-shell is in this legion always simultaneously developed (suddenly excreted at the moment of lorication from the sarcodictyum). The secondary lattice-shells, on the other hand, which surround the former concentrically, and are united with it by radial bars, arise successively from within outwards.
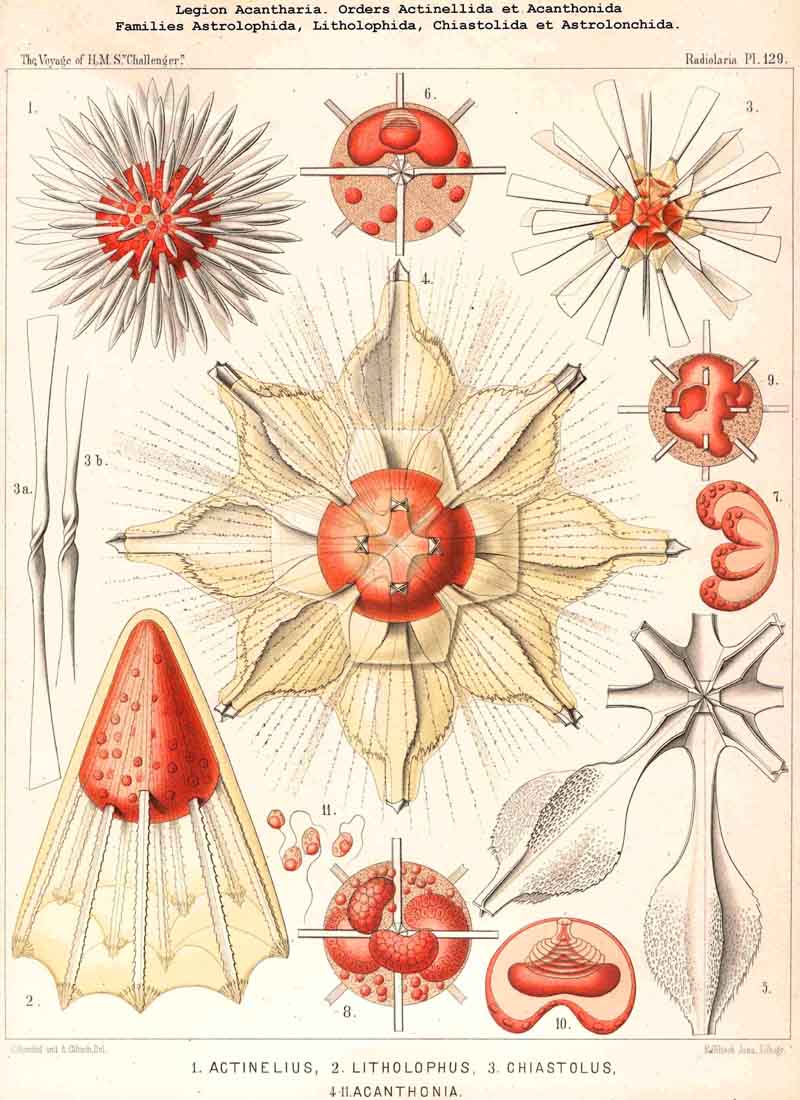
147. The Ontogeny of the Acantharia.-The individual development of the ACANTHARIA in the simplest case (Actinelius) stops at a point which differs from the Actissa-stage only in the change of radial axial threads into acanthin spines. In the small group Actinelida, their number remains variable and usually indeterminate (Adelacantha), whilst in the great majority of the legion (Acanthonida and Acanthophracta) the number is constantly twenty, and those spines are regularly arranged according to the Müllerian law in five parallel circles, each containing four crossed spines (Icosacantha). The simplest form among these latter is Acanthometron, which may be regarded both ontogenetically and phylogenetically as the common starting-point of all the Icosacantha. Within this extensive group variations in the length of the dimensive axes appear, similar to those observed in the SPUMELLARIA. In the Astrolonchida and Sphaerophracta the central capsule remains spherical, extending equally in all directions ; and correspondingly the lattice-shell, which is excreted on the surface of the spherical calymma, remains spherical. In the Belonaspida (just as in the Prunoidea) this form passes over into an ellipsoid by prolongation of one axis ; on the contrary, in the Hexalaspida (as in the Discoidea) the discoidal or lenticular form arises by shortening of an axis. Finally, in the Diploconida, and in some Hexalaspida in which the growth is different in all three dimensive axes (as in the Larcoidea), both the central capsule and the shell assume the lentelliptical form. The lattice-shell of the Acanthophracta is usually successive in its development, since from each of the twenty radial spines two or four tangential apophyses proceed, whose branches subsequently unite and combine to form the lattice-shell. Only in the peculiar Sphaerocapsida can the pavement-like shell arise simultaneously or in a moment of lorication.
148. The Ontogeny of the Nassellaria.-The individual development of the NASSELLARIA in the simplest instance remains stationary at the skeletonless Nasselid stage (Cystidium, Nassella), which can be immediately derived from the foregoing Actissa stage by the disappearance of the pores in the upper (apical) hemisphere of the central capsule, whilst in the lower (basal) portion they are modified to form a porochora ; the podoconus is developed within the endoplasm upon this latter. Usually the spherical form of the central capsule passes over into an ovoid or ellipsoidal one, the vertical axis which passes through the centre of the porochora being elongated. From the skeletonless Nassellida the other NASSELLARIA may be derived both ontogenetically and phylogenetically by the excretion of an extracapsular siliceous skeleton. Unfortunately, the earliest stages in the formation of this skeleton are unknown, and hence no answer can at present be given to the important question, in what order the three primary skeletal elements of the NASSELLARIA (the basal tripod, sagittal ring, and latticed cephahis) appear (compare §§ 111 and 182). If, for example, in Cortina and Tripospyris the basal tripod were to appear first in the ontogeny, and the sagittal ring were developed from this, then the Plectoidea would be rightly considered to be the oldest forms in the phylogeny of the skeleton-forming NASSELLARIA ; and in the contrary case the Stephoidea would be so regarded. The relations of growth in the three dimensive axes are very variable in the NASSELLARIA ; the three most important factors in this respect (partly separately and partly in combination) are ; (1) the development of the basal tripod to a triradial stauraxon form (the ground-form being a three-sided pyramid) ; (2) the development of the sagittal ring in the median plane of the body (the vertical axis having the poles different) ; (3) the development of the latticed cephalis outside the central capsule (the poles of the vertical axis being again different). Since the development both of the skeleton and of the malacoma is characterised in most NASSELLARIA by the stronger growth of the vertical axis and the differentiation of the two poles, the allopolar monaxon ground-form acquires a predominant significance in this legion (§ 32) ; the starting point of most of the further modifications is the basal pole of the vertical main axis. Next to this the sagittal axis is usually the most important determining factor (its dorsal and ventral poles being usually different), more rarely the frontal axis (with equal right and left poles). In the zygothalamous Spyroidea (as in the Stephoidea) the formation of the shell proceeds from the sagittal ring, whilst in the polythalamous Cyrtoidea the latticed cephahis is always the starting point, from which a series of joints (thorax, abdomen, and in the Stichocyrtida, the numerous post-abdominal joints) successively arise (unipolar growth).
149. The Ontogeny of the Phaeodaria.-The individual development of the PHAEODARIA in the simplest case stops with the skeletonless condition of the Phaeodinida (Phaeodina, Phaeocolla), which can be immediately derived from the foregoing Actissa-stage by the disappearance of the pores in the greater part of the central capsule, the characteristic astropyle being developed at the basal pole (§ 60). Since this particular form and structure of the spheroidal central capsule remains the same in all PHAEODARIA, whilst the formation of their skeleton follows very different directions, it follows that further common paths of development are excluded both ontogenetically and phylogenetically. What will be laid down in this respect as regards the phybogeny of the different groups of PHAEODARIA (§§ 194-199) holds true also of their ontogeny. The relations of growth In the three dimensive axes are hence very different in the skeletons of the various groups of PHAEODARIA. This difference is best marked in the Phaeoconchia, whose bivalved lattice-shells have as their ground-form the rhomboid pyramid of Ctenophora. In most Phaeogromia the monaxon lattice-shell may develop simultaneously by sudden excretion at a particular moment of lorication ; this is also the case with the polyaxon lattice-shells of the Phaeosphaeria. In their further growth the develop ment of basal or radial apophyses is of special importance. In the majority of the PHAEODARIA these apophyses are tubes of silicate filled with jelly (often provided with an axial siliceous thread) ; thus their development is distinguished by complications which are absent in the case of the other three legions.
150. Growth.-The growth of the body in the Radiolaria, as in all other organisms, is the fundamental function of individual development (see note A). All structural relations which this richest class of the Protista exhibits may be referred to different forms of growth, either of the unicellular malacoma or of the skeleton which it produces. In general the special development of the skeleton is dependent upon that of the central capsule, and of the sarcodictyum on the surface of the calymma ; in the further growth, however, the conditions are reversed, and the condition of the skeleton already formed directly determines the further development of the central capsule and of the calymma with its sarcodictyum. The four legions of Radiolaria show, speaking generally, certain characteristic differences in growth, which are due in great part to the different structure and ground-form of their central capsule. In the two legions of the Porulosa (SPUMELLARIA and ACANTHARIA), in which the central capsule is originally spherical and the ground-form of the skeleton either polyaxon or isopolar monaxon, two fundamental and variously combined directions of growth are recognisabbe ; firstly, the concentric growth (equal increase of volume in all directions), and secondly, multipolar or diametral growth (hypertrophy of certain parts in the direction of definite pairs of radii). A different state of things obtains, however, for the most part, in the two legions of the Osculosa (NASSELLARIA and PHAEODARIA), in which the central capsule possesses a vertical main axis with different poles, and the structure of the skeleton is determined by this allopolar monaxon ground-form. The two fundamental directions of growth here combined in the most various ways are, firstly, unipolar growth (starting from the basal pole of the vertical main axis), and secondly, radial or pyramidal growth (characterised by the different development of separate parts in the direction of definite radii). Whilst the growth of the malacorna is dependent on intussusception (as in most organic structures capable of imbibing), the growth of the skeleton in all Radiolaria takes place by apposition (see note B).
A. The earliest investigations into the modes of growth in the Radiolaria are due to J. Müller (L. N. 12, pp. 21 -33). More detailed communications I gave myself in my Monograph (L. N. 16, pp. 150-159). The relations there sketched have now, in consequence of the examination of the Challenger collection, undergone many important additions, and in some divisions, important modifications ; these are for the most part treated of in the general account of the separate families.
B. The view here maintained, that the skeleton of all Radiolaria grows only by apposition, appeared formerly to have certain exceptions. I thought I had shown that in Caelodendrum the thin-walled tubes grew not only in length but also in thickness, with continuous increase in the lumen (L. N. 16, pp. 152, 360). Further K. Brandt concluded, from the varying size of the median bars in the twin-spicules of Sphaerozoum, that these siliceous structures grow by intussusception (L. N. 38, p. 401). Both suppositions have been proved erroneous, and I have come to the opinion that in all Radiolaria the skeleton grows by apposition.
151. Reqeneration.-Whilst the general course of individual development (perhaps without any exception in the Radiolaria), begins with the formation of zoospores in the central capsule, there yet occurs in some groups a different form of ontogeny, introduced by simple division of the unicellular organism, and coming under the term "regeneration" in its wider sense. This spontaneous division occurs quite commonly in the Polycyttaria (or social SPUMELLARIA), and produces their colonies (compare the chapter on Reproduction, § 273). On the contrary, it has not been observed in the solitary SPUMELLARIA, nor in the ACANTHARIA and NASSELLARIA ; possibly, however, the peculiar ACANTHARIAN family, Litholophida, has arisen by the division of Acanthonida (compare p. 734). Among the PHAEODARIA division is commonly observed in the order Phaeocystina (which have an incomplete Beloid skeleton or none), and also in the Phaeoconchia. In all these cases the increase by division is nothing else than an ordinary case of cell-division, in which bisection of the nucleus precedes that of the central capsule. The regeneration by which each of the two daughter-cells develops to a complete mother-cell depends upon simple growth. Another form of regeneration, different from this, has been observed in Thalassicolla. If the central capsule be extracted artificially from the large concentric calymma, the enucleated central capsule produces a new extracapsulum, with sarcomatrix, pseudopodia, and calymma. This experiment may be repeated several times with the same result. (Compare A. Schneider, 1867, L. N. 20.)
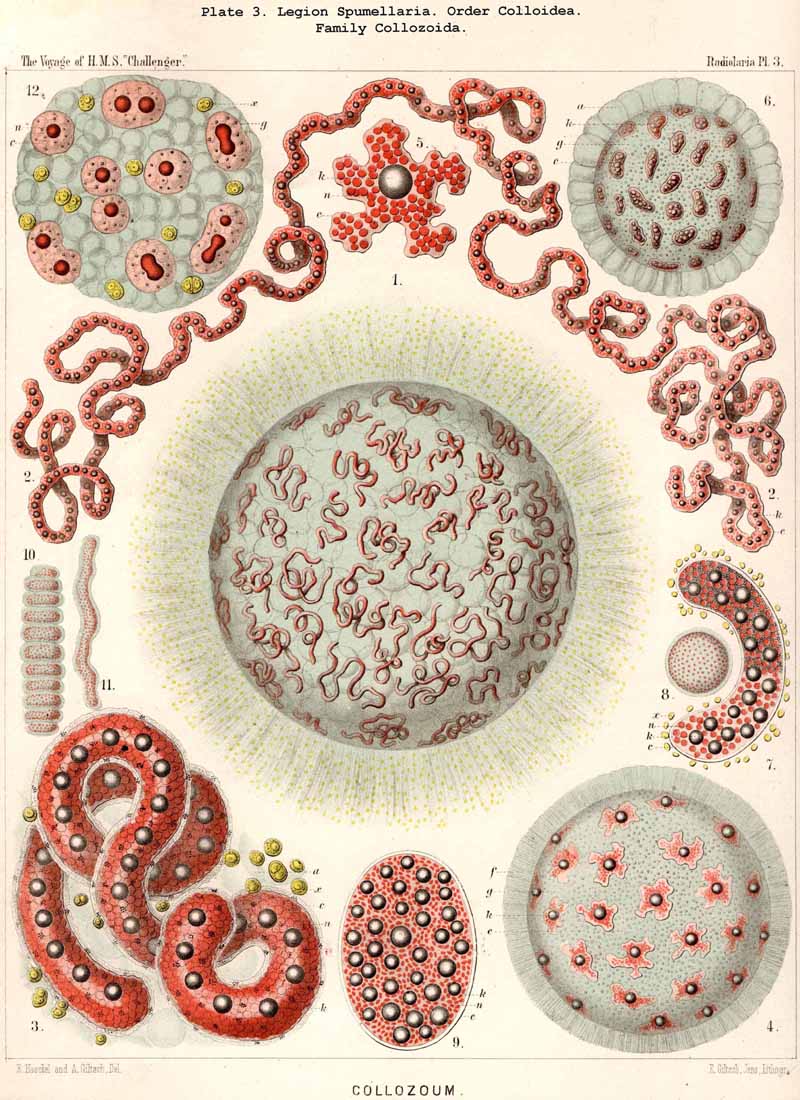
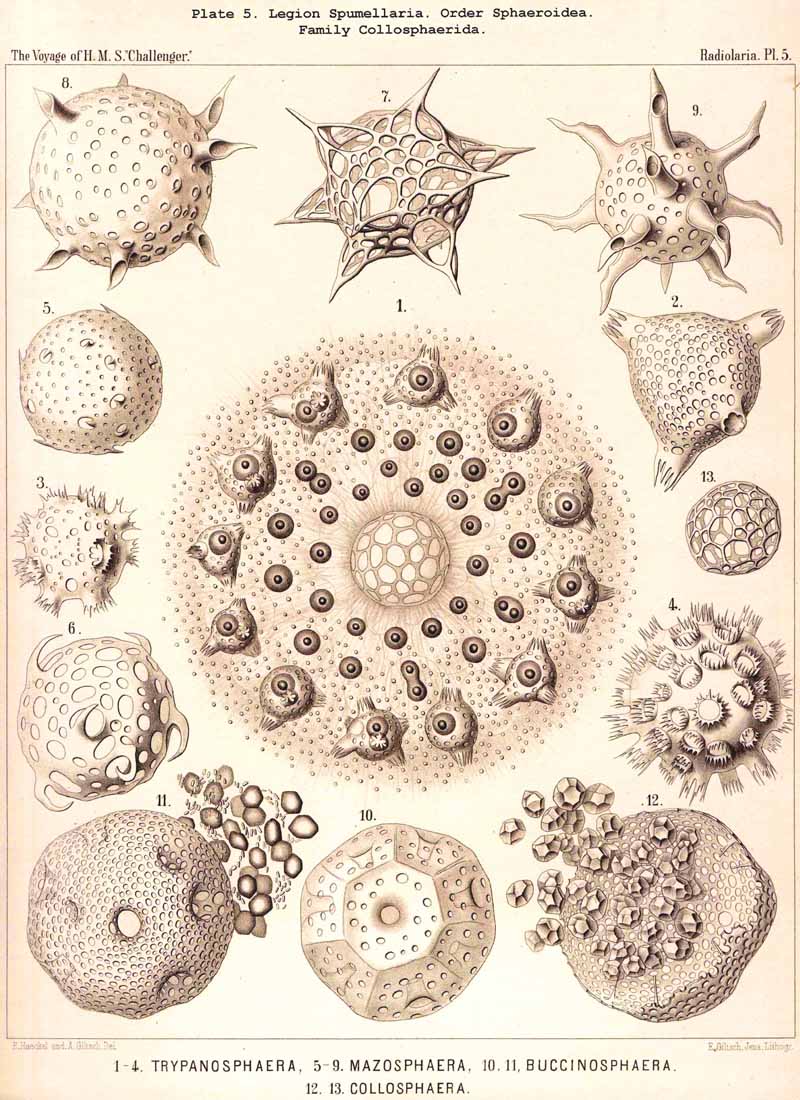
152. The Formation of Colonies.-The individual development of colonies takes place in all three families of the Polycyttaria (Collozoida, Sphaerozoida, Collosphaerida) in the same simple way, by the repeated division of a single monozootic SPUMELLARIAN. Since these divisions only affect the central capsule and not the extracapsulum, the sister-cells, which arise by repeated division of the mother, remain enclosed in a common rapidly growing calymma. Probably in all Polycyttaria the commencement of the formation of colonies immediately follows the Actissa-stage of the monozootic mother-cell (or takes place in the Thalassicolla.-stage, which arises from the former by the development of alveoles in the calymma). The simple central nucleus separates (by direct nuclear division) into two halves, and the central capsule follows this process of bisection, becoming constricted in the middle between the two daughter nuclei (Pl. 3, fig. 12). In the further growth of the colony the process of division proceeds in the older, now multinucheate, central capsules, in which an oil-globule has taken the place of the original nucleus ; then the division of the oil-gbobules precedes that of the central capsule (Pl. 5, fig. 1). Another mode of growth of the colonies is the multiplication of the central capsules by gemmulation, or the formation of the so-called "extracapsular bodies" (Gemmulae, § 214). The characteristic skeletal structure of the different species appears at a later stage. Whether ripe central capsules can emerge from the social bond of a coenobium, and, having become isolated, establish the formation of a new colony, is very doubtful. The various forms which the coenobium assumes in the different species of Polycyttaria, are due partly to simple growth, partly to the development of large vacuoles in the calymma.
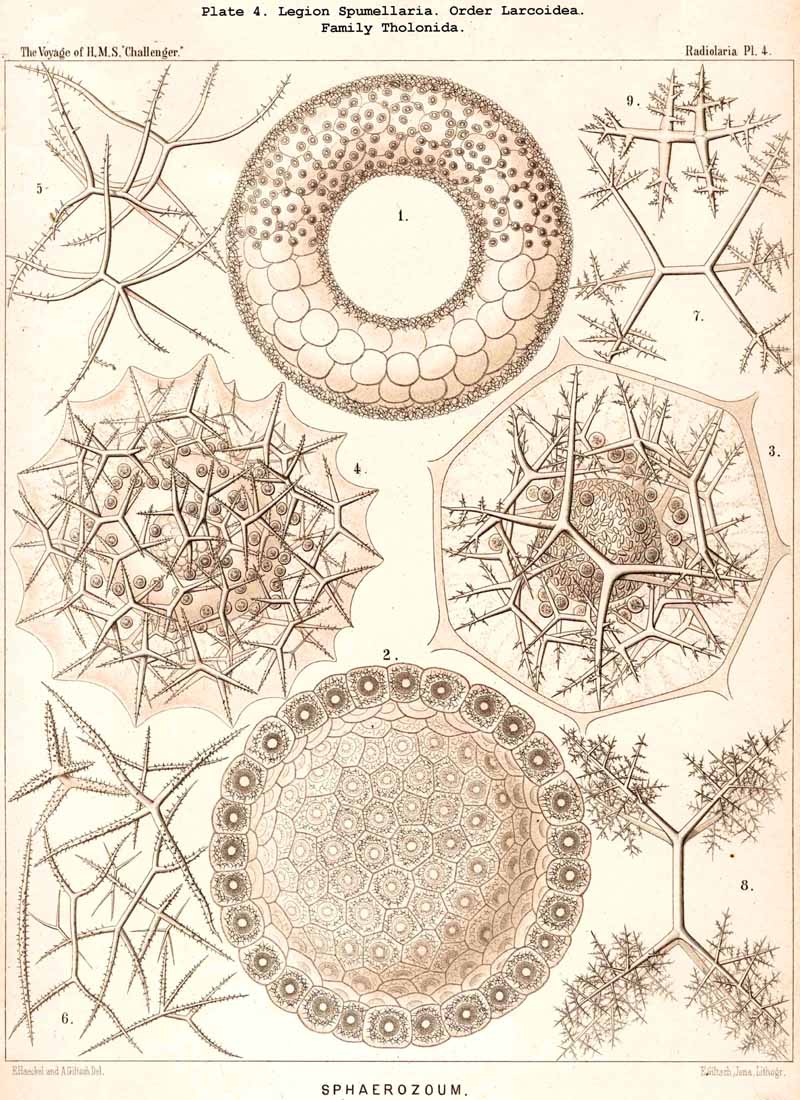
The form and size of the coenobia appear in many fully developed Polycyttaria to exhibit specific differences, which require further investigation ; in the young stage, on the contrary, they are simple spheres or ellipsoids, often cylindrical or sausage-shaped (Pl. 3, figs. 1, 4, 6, 11). In some species the cylindrical gelatinous bodies become moniliform, and separated by transverse constrictions into many segments, each of which encloses a large alveole (Pl. 3, fig. 10). The rare ring-shape (Pl. 4, fig. 1) which I figured in 1862 in the case of Collozoum (L. N. 16, p. 522, Taf. xxxv. fig.1), I have recently observed in different species of Polycyttaria ; it is capable of a very simple mechanical explanation, both ends of a sausage-shaped colony having been accidentally brought into contact by a wave and having united by agglutination. Quite recently Brandt has given a very complete account of the development, form, and growth of Polycyttarian colonies in his work on the colonial Radiolaria of the Bay of Naples (1885, L N. 52, pp. 71-85).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}